Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
C. elegans vulval development is an intensively studied example of animal organogenesis. A network of intercellular signaling, signal transduction, and transcriptional regulation underlies the precise formation of this organ, which is the connection between the hermaphrodite uterus and the outside of the nematode. A single cell of the somatic gonad, the anchor cell, organizes the development of the vulva from epidermal precursors as well as the physical connection of the epidermis with the uterus. WNT signaling acting via the HOX gene lin-39 renders six epidermal precursor cells competent to respond to other developmental signals. The anchor cell induces nearby epidermal precursor cells to generate vulval cells via an epidermal growth factor (EGF) signaling pathway. The precise pattern of vulval precursor cell fates involves the graded action of the EGF signaling and LIN-12 (Notch) mediated lateral signaling. EGF promotes the 1° fate while LIN-12 promotes the 2° fate. Both EGF and LIN-12 prevent precursor cells from adopting the 3° fate, which generates non-specialized epidermis. EGF-receptor and Notch signaling are antagonistic: EGF-receptor signaling leads to down-regulation of the Notch-like receptor LIN-12, while LIN-12 signaling induces negative regulators of EGF-receptor signaling such as MAP kinase phosphatase LIP-1 and the tyrosine kinase ARK-1. The 1° precursor cell generates vulE and vulF mature vulval cells; the pattern of vulE and vulF cells requires an additional signal from the anchor cell as well as WNT signaling. The two 2° precursor cells generate vulA, vulB1, vulB2, vulC and vulD cells but in mirror symmetric polar patterns: ABCD and DCBA. The reversed polarity of the posterior 2° precursor cell lineage requires WNT signaling mediated by both Frizzled class and Ryk class WNT-receptors LIN-17 and LIN-18, respectively. A network of transcription factors controls the seven mature adult cell types; these include the LIM domain protein LIN-11, the Pax2/5/8 protein EGL-38, the zinc finger protein LIN-29, and the Nkx6.1/6.2 protein COG-1. The anchor cell also patterns nearby uterine cells, via the DSL ligand LAG-2 and LIN-12, to generate the four uv1 cells that form the tight connection with the vulva. This connection is initiated by the anchor cell, which invades between the vulF cells in a process analogous to invasive behavior of metastatic tumor cells. During this invasion process, the basement membranes between the gonad and body wall are degraded. The extensive information about vulval development in C. elegans has helped it become a paradigmatic case for identifying and studying a variety of regulatory pathways.
The C. elegans hermaphrodite vulva develops during postembryonic (larval) development from ventral epidermal precursors, and connects the developing uterus to the external environment. In the adult, the vulva is necessary for egg-laying (see Egg-laying) and for copulation with males (see Male mating behavior). Vulval development has attracted general interest for three main reasons. First, it serves as a paradigm for organogenesis. In particular, vulva development represents a well-understood case in which invariant development arises from multiple cell-cell interactions. It is also a striking example of tissue remodeling: the formation of a hole at a precise location in an organism. Second, it has been important for the genetic analyses of signaling and signal transduction by epidermal growth factor (EGF)-receptor LET-23 and RAS LET-60; (see RTKRas/MAP kinase signaling), LIN-12 (see LIN-12/Notch signaling in C. elegans), and WNT (see Wnt signaling), as well as the functions of the SynMuv and mediator proteins (see Transcriptional regulation). Third, it has become a paradigm for examining the interactions among regulatory pathways, notably the antagonism of EGF-receptor (LET-23) and Notch (LIN-12) pathways.
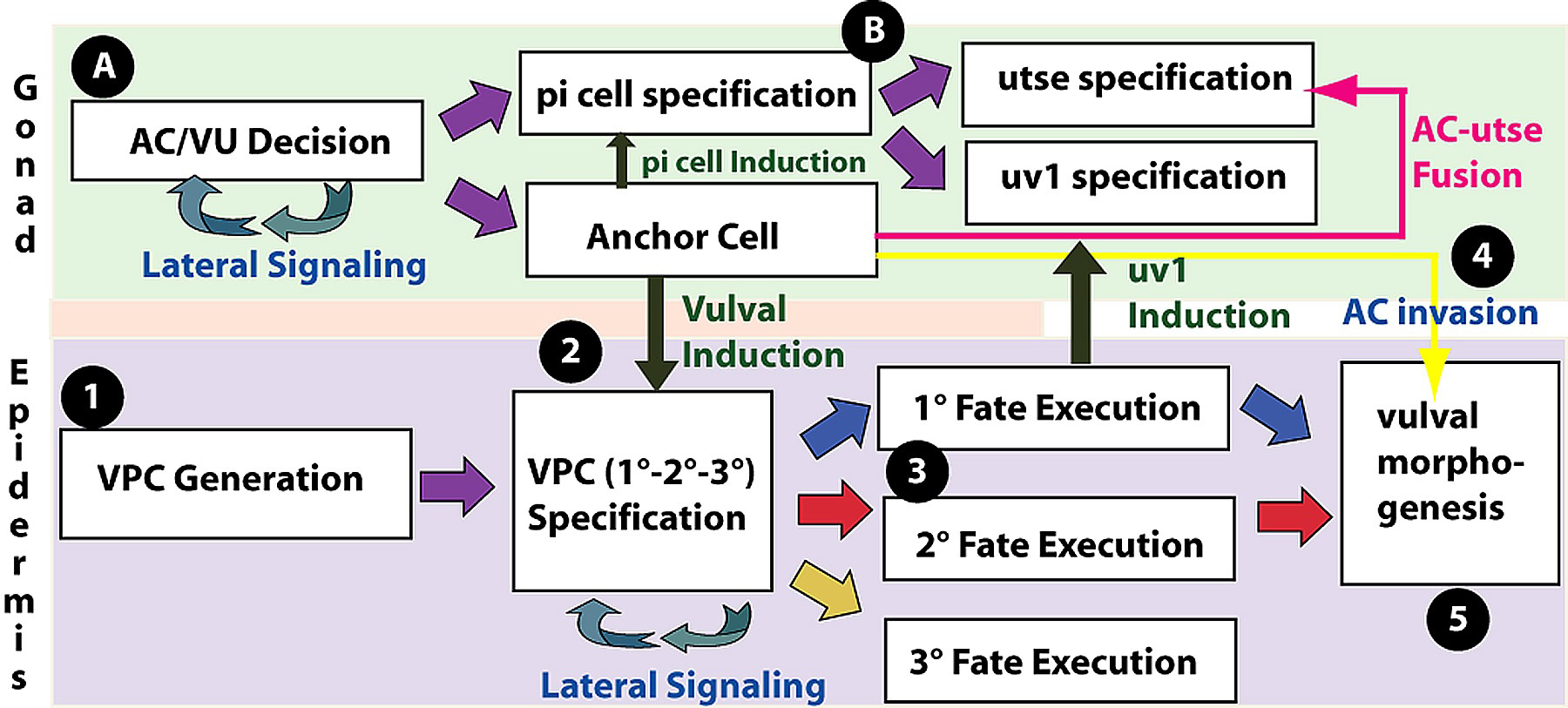
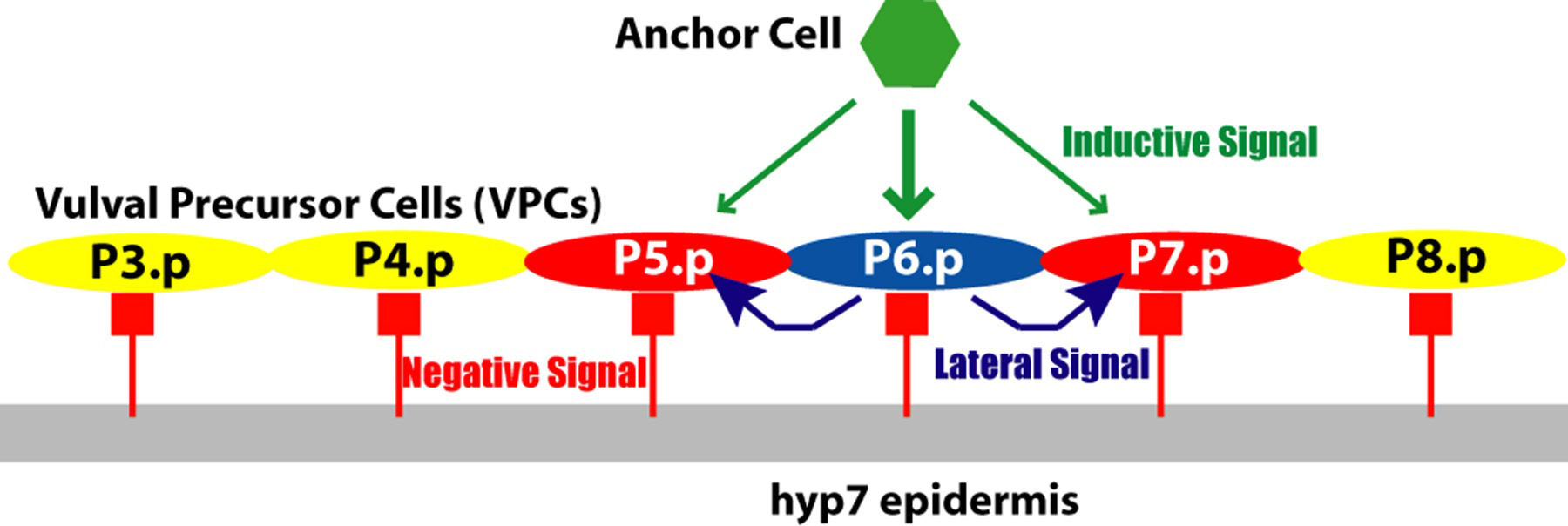
Vulval development is conveniently thought of as occurring in five steps (Figure 1). (1) Generation of VPCs: During the L1 and L2 stages, six vulval precursor cells (VPCs) are specified among the 11 Pn.p cells, which are located in the ventral epidermis. (2) Vulval Precursor Patterning: During the L3 stage, a signal from the gonad and signaling among the VPCs specifies three VPCs to generate vulval cells (Figure 2). The vulval lineages are of two types, 1° and 2°, each of which generate distinct sets of progeny. The uninduced VPCs generate a 3° lineage, which make epidermal cells that fuse with the large syncytial epidermis hyp7. In an intact, wild-type hermaphrodite, the VPCs adopt their fates in a precise spatial pattern: 3°-3°-2°-1°-2°-3° (Figure 3). (3) Generation of the adult cells: The adult vulva comprises 22 nuclei in cells of seven types, vulA, vulB1, vulB2, vulC, vulD, vulE, and vulF, that differ in their patterns of gene expression and in details of their cell biology. (4) Anchor cell invasion: the anchor cell extends process to the center of the vulF cells and forms a hole in the epidermis. (5) Morphogenesis of the vulva: the seven types of vulval cells invaginate, and sequentially form seven distinct toroids, connects to the uterus, and everts as the hermaphrodite molts to adulthood. In addition, two aspects of uterine development are crucial to the development of the vulva. (A) Generation of the anchor cell. The anchor cell is specified from among two somatic gonadal cells during the end of the L2 stage (see AC/VU decision). (B) Uterine Patterning. The anchor cell also patterns the developing uterus, inducing the six pi cells, which generate the utse cell and uv1 cells. The uv1 cells attach to the vulF cells, while the anchor cell ultimately fuses with the utse.
|
Figure 1. Vulval development is a multi-step process. It involves the coordinated development of the uterine (green, top) and ventral epithelia (purple, bottom) with several interactions between these tissues. 1. The six VPCs are specified from among the ventral epidermal Pn.p cells. 2. The VPCs become specified to 1°, 2° or 3° cell fates by multiple signaling pathways. The anchor cell induces vulval fates (1° and 2°) and lateral signaling among the induced VPCs specifies the 2° VPC fate. 3. The vulval lineages are generated and specific sets of mature cells are produced and specified. 4. The anchor cell breaks down the basement membrane of the gonad and vulval epithelia and invades between the vulF cells. 5. The vulval cells undergo a complex morphogenesis.A. The anchor cell becomes specified from a set of two AC/VU cells; the cell that is not the anchor cell becomes one of three VU precursor cells. B. The three VU cells generate 12 grandprogeny, six of which are induced by the anchor cell to become pi cells. The six pi cells divide, and four of their progeny are induced by the vulF cells of the vulva 1° lineage to become uv1 cells. The remaining eight cells fuse to form the utse, which ultimately fuses with the anchor cell.
|
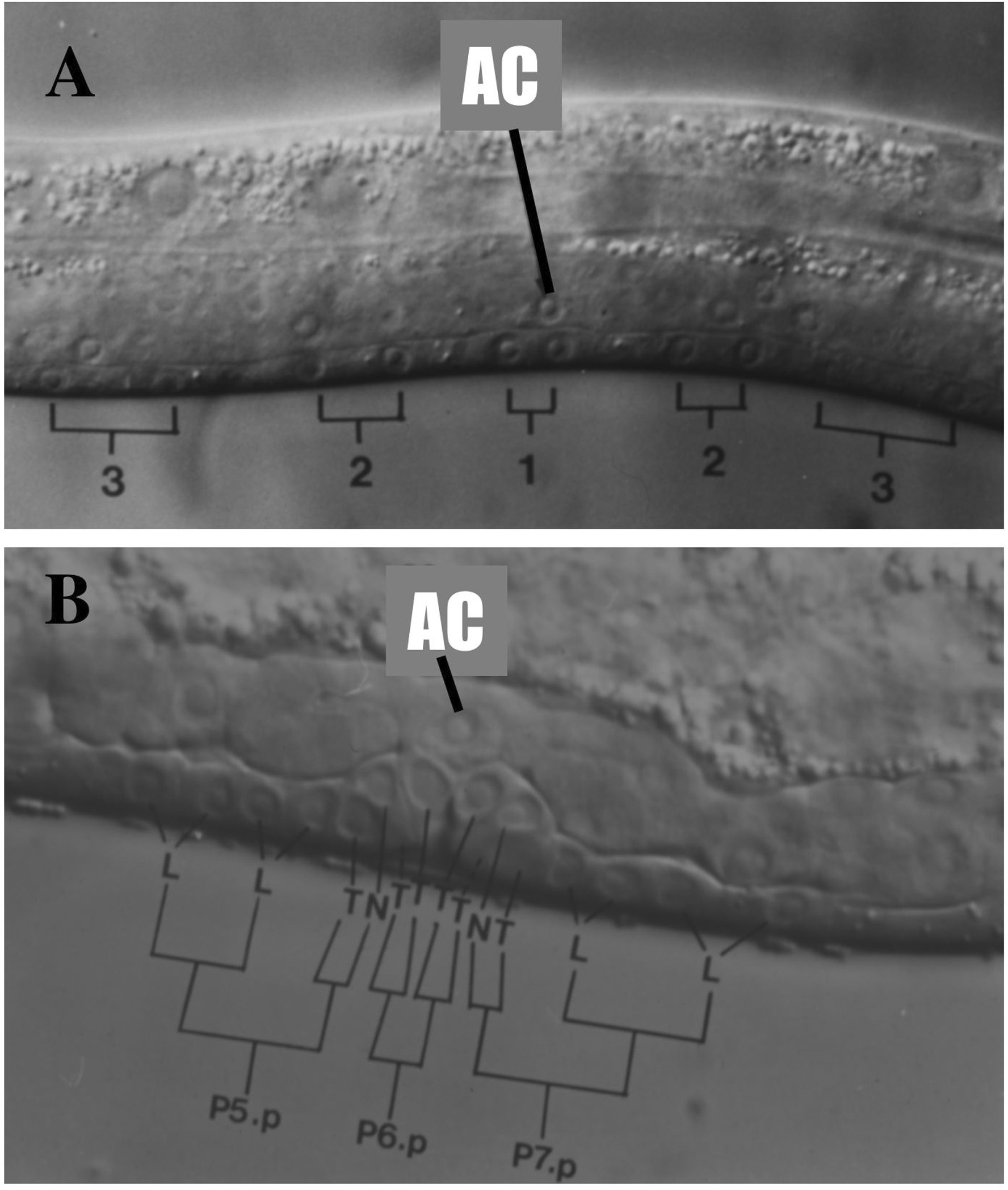
Figure 2. Nomarski images of developing vulva. Left lateral views of larval hermaphrodites (anterior to the left). AC, anchor cell. 3, 3° VPCs (P4.p and P8.p shown here); 2, 2° VPCs (P5.p and P7.p); 1, 1° VPC. (A) mid-L3 stage . At this stage, there are no signs of morphological differentiation but P6.p and its daughters express egl-17. B. L3 lethargus. The first two rounds of mitosis of the 2° and 1° vulval precursors are drawn in, with the axis of the third round of division indicated: L, longitudinal (anterior-posterior); T, transverse (left-right); N, no division; S, fuses with hyp7 epidermal syncytium. In the literature, 1° lineages are referred to as [TTTT] or [EFFE]; 2° lineages are referred to as [LLTN], [NTLL], [ABCD], or [DCBA]; 3° lineages as [S S]; hybrid lineages are [S TN], [NT S], [S TT], [TT S], [LL S] or [S LL]. In abnormal lineages, O refers to an oblique axis of division, and U refers to undivided cell.
|
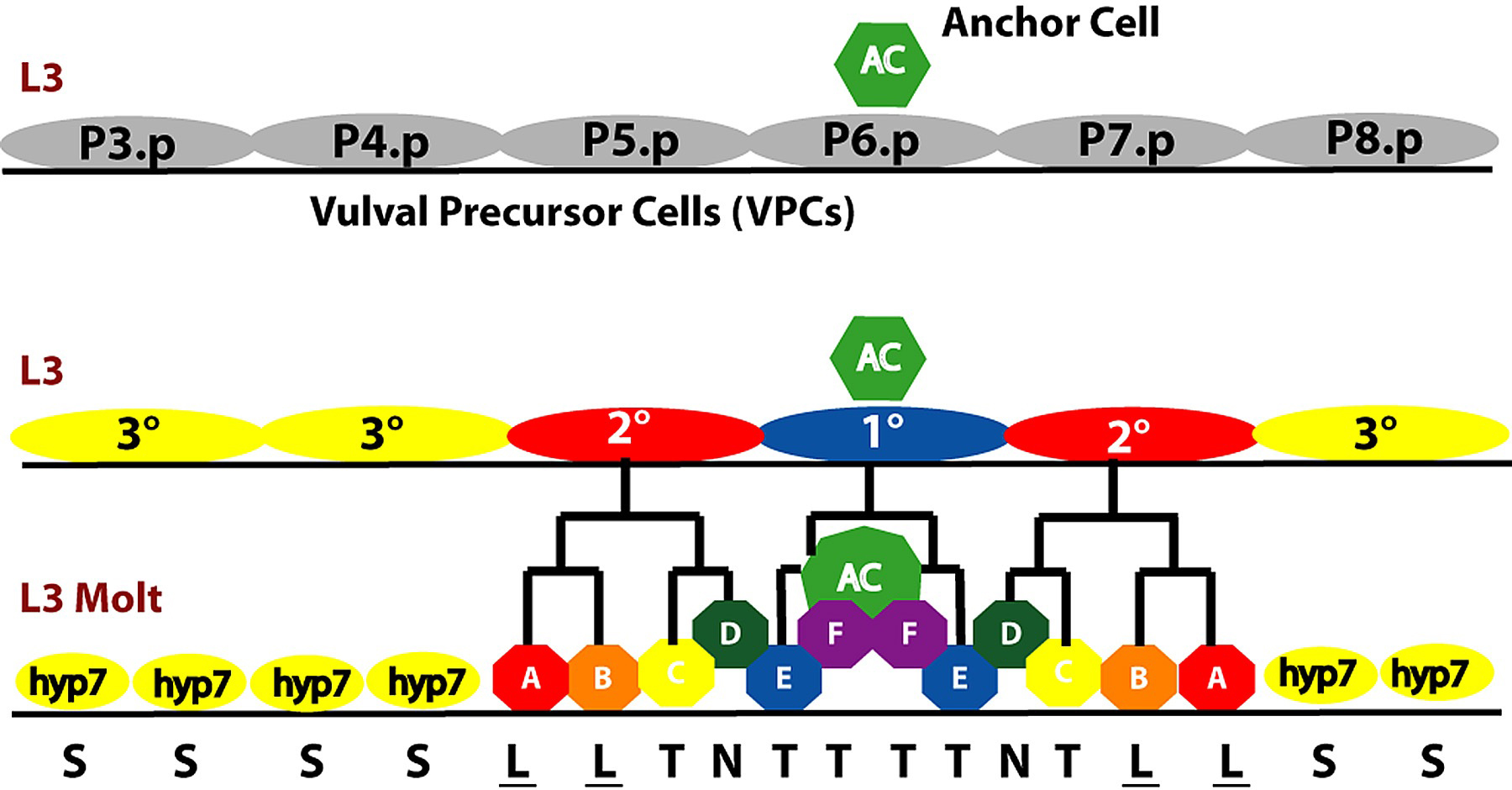
Figure 3. Simple view of vulval lineages. The six multipotent VPCs become specified during the early L3 stage to an invariant pattern of VPC fates. The 1° and 2° VPCs generate distinct sets of progeny cells. Hyp7, cell fuses with the large epidermal syncytium hyp7. A, generates two vulA nuclei; B, generates a vul B1 and a vulB2 cell; C, generates two vulC cells; D, becomes vulD; E, divides to generate two vulE cells; F, generates two vulF cells. Most of the VPC granddaughters shown L, longitudinal axis of cell division; T, transverse axis of cell division; N, no cell division. Underline indicates adherence to the ventral cuticle. See Sulston and Horvitz (1977) for lineages and Katz et al. (1995) for morphological distinctions between 1° and 2° lineages.
Mutations that affect vulva development often cause defects that are visible in the dissecting microscope. These phenotypes include the inability of hermaphrodites to lay eggs (EGg-Laying-defective; Egl), or morphological defects such as Protruding VuLva (Pvl) or abnormal Eversion of the vuLva (Evl). Specifically, mutants with defective vulval induction, e.g., mutations that decrease activity of the EGF-receptor let-23 (see below) cause a VULvaless (Vul) phenotype. Vulvaless hermaphrodites have fertile eggs that hatch internally and eventually escape from the corpse of their mother. Hermaphrodites with defects in the vulval lineages or in the vulval-uterine connection have protruding vulvae or are egg-laying defective. Mutations that activate LET-23, rendering it constitutive, result in a MUltiVulva (Muv) phenotype. Multivulva hermaphrodites typically have a single functional vulva and additional ventral protrusions, each a pseudovulva formed from vulval tissue. Mutants with reversed polarity of P7.p 2° lineage have a psuedovulva posterior to the normal vulva.
In addition to screens for mutants with vulval defects (Horvitz and Sulston, 1980; Ferguson and Horvitz, 1985; Seydoux and Greenwald, 1993; Eisenmann and Kim, 2000; Palmer et al., 2002), suppressor screens proved useful in identifying new genes and alleles, including members of the LET-23 pathways and genes involved in other steps in vulval development (e.g., Beitel et al., 1990; Han et al., 1990; Aroian et al., 1991; Clark et al., 1992; Clark et al., 1992; 1993; Han et al., 1993; Clark, 1992).
The phenotype of a hermaphrodite observed at the dissecting microscope level can sometimes be misleading, and thus the dissecting microscope phenotypes should always be confirmed by observation of anatomy in a compound microscope. For example, lin-17 mutants often display a vulva (from P5.p and P6.p) and a posterior pseudo-vulva (from P7.p), appearing to be multivulva, but have a normal extent of vulval induction. A number of markers are now available to help distinguish different fates of VPCs (Burdine et al., 1998; Yoo et al., 2004) and of terminal vulval cells (Inoue et al., 2002).
The eleven Pn.p cells (P1.p, P2.p, P3.p,..., P11.p) are formed during the L1 larval stage as the posterior daughters of the Pn ventral neuro-ectoblasts (P1, P2, P3, ...., P11), the anterior daughters of which generate primarily ventral cord neurons (Sulston and Horvitz, 1977). The Pn cell nuclei migrate into the ventral cord during the L1 stage. The development of the vulva thus depends upon the generation of the Pn.p cells. Action of the Hox genes is responsible for specification of the VPCs P3.p, P4.p, ..., P8.p (P(3-8).p; Figure 4). In particular, lin-39 is necessary for specification of the VPCs; in the absence of lin-39 function, P(3-8).p fuse with hyp7 epidermis in the L1 larval stage (Salser et al., 1993; Clark et al., 1993; Figure 5). In wild-type hermaphrodites, P3.p sometimes fuses with hyp7 during the L2 stage (Sulston and Horvitz, 1977).
|
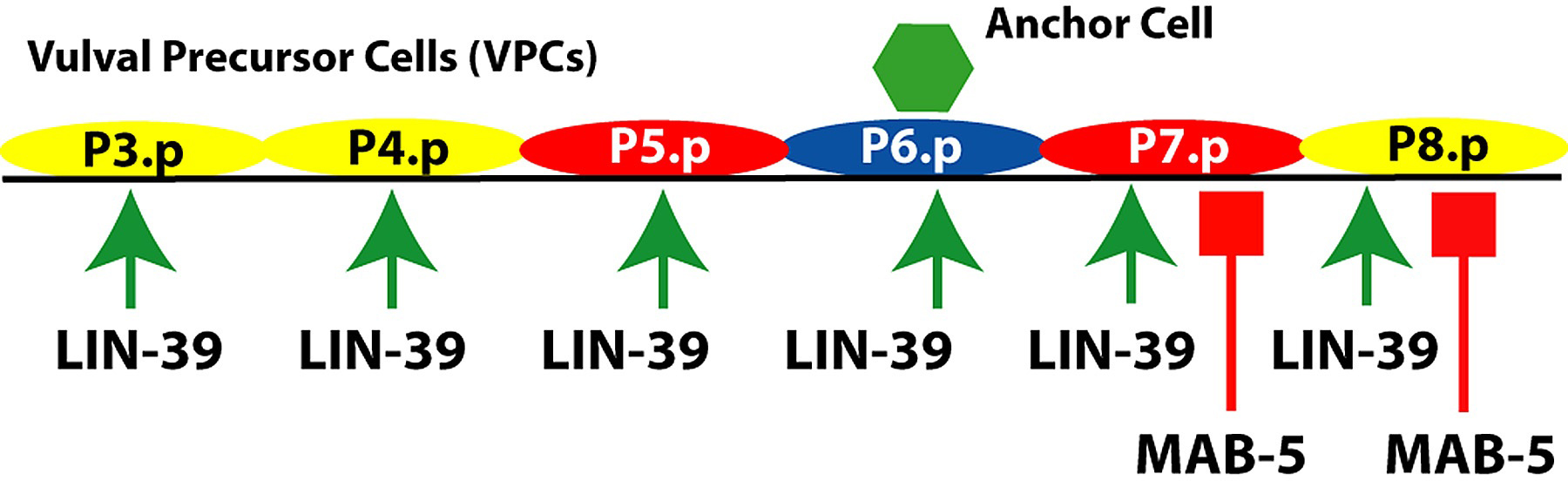
Figure 4. Hox genes and VPC competence group. lin-39 is expressed in the central body region, including P3.p- P8.p shown. mab-5 is expressed in more posterior cells including P7.p and P8.p. Arrow, positive regulation of competence; bar, negative regulation of competence. Yellow, 3° fate; red, 2° fate; blue, 1° fate.
|
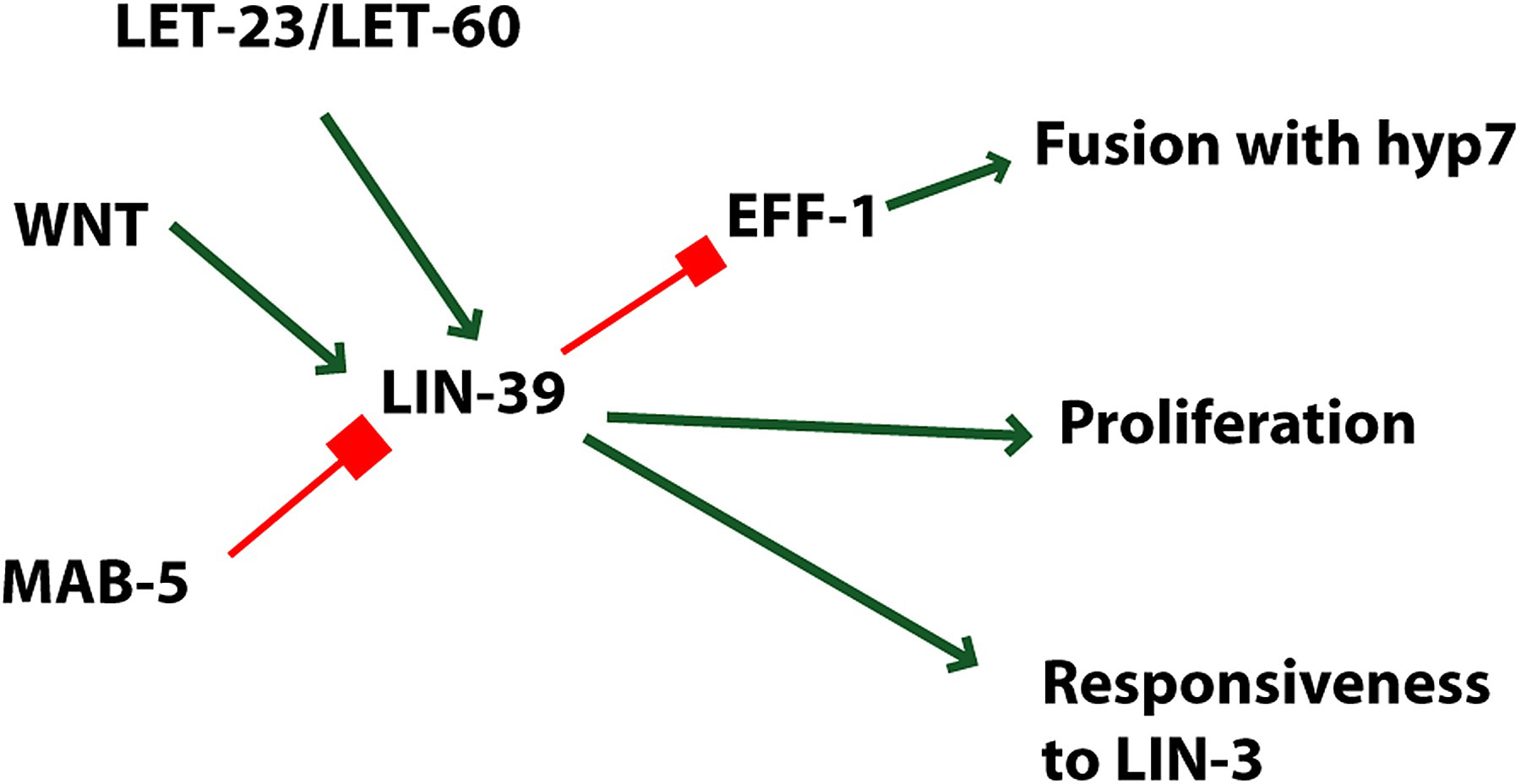
Figure 5. Roles of LIN-39. lin-39 is positively regulated by WNT and LET-23 signaling, and is proposed to be inhibited by MAB-5 (see Figure 3). LIN-39 regulates proliferation, responsiveness to LIN-3 and inhibits fusion with hyp7 via inhibition of EFF-1 expression. These events are occurring in the VPCs; transduction of the Wnt signal occurs in the VPCs, but the source of the Wnt signal is unknown. Green arrows, positive regulation; red bars, negative regulation.
In the intact, wild-type hermaphrodite grown under standard conditions, P5.p, P6.p and P7.p generate vulval cells. Ablation experiments indicated that the six cells P3.p, P4.p, P5.p, P6.p, P7.p and P8.p are competent to respond to intercellular signals and generate vulva. (Sulston and White, 1980; Kimble, 1981; Sternberg and Horvitz, 1986). For example, ablation of P6.p may result in P5.p adopting the 1° fate and P4.p adopting the 2° fate, or, P7.p may adopt the 1° fate and P8.p the 2° fate. Ablation of P5.p may result in replacement by P4.p, whle ablation of P7.p may result in replacement by P8.p. Ablation of all VPCs but P3.p results in P3.p becoming 1°. There is some variability, especially as a function of time at which the ablation occurs.
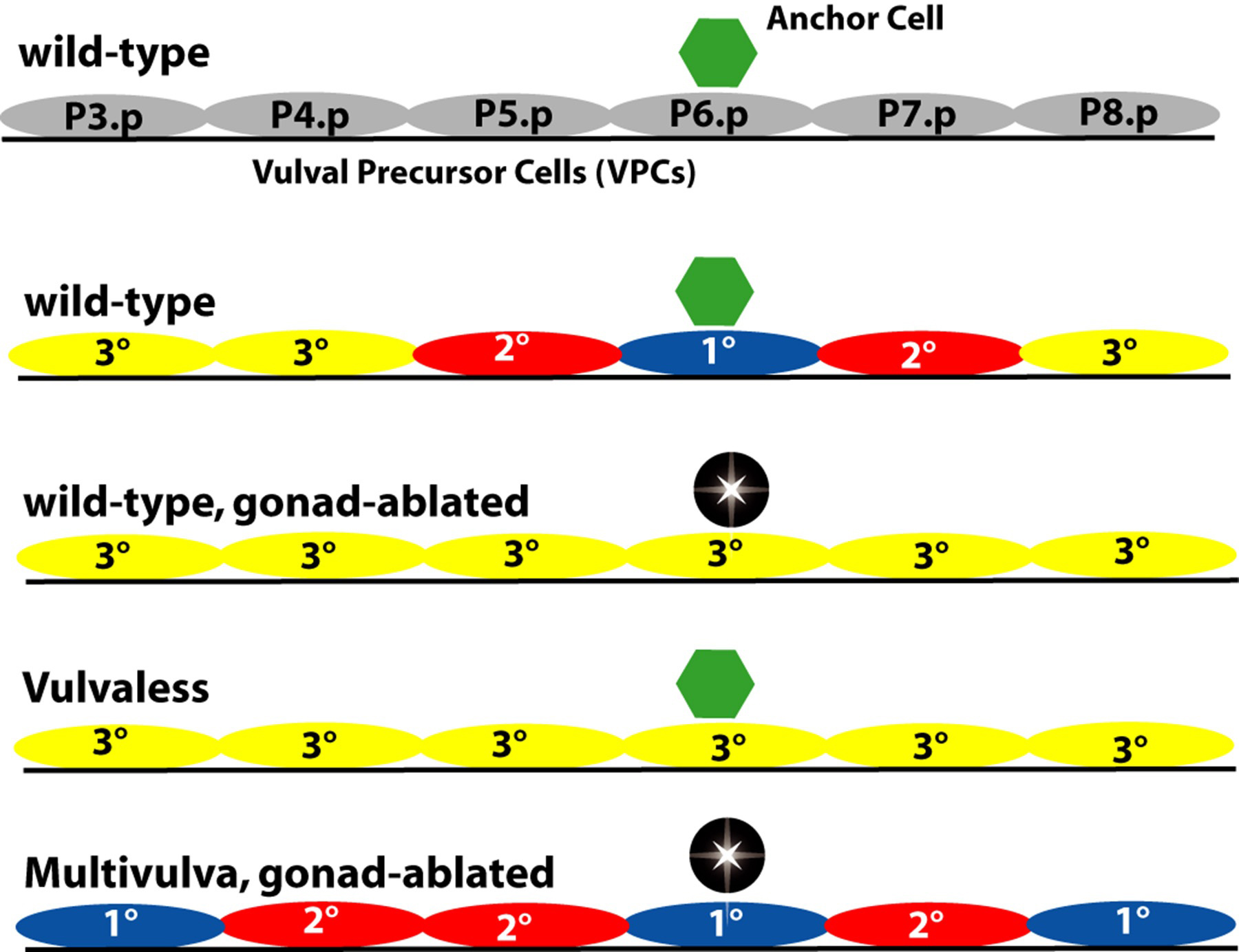
Ablation of the gonad during the L1 stage (Sulston and White, 1980), or of the anchor cell prior to the L3 stage, results in all six VPCs becoming 3° (Kimble, 1981; Figure 6). These experiments also indicate that the anchor cell induces the vulva.
|
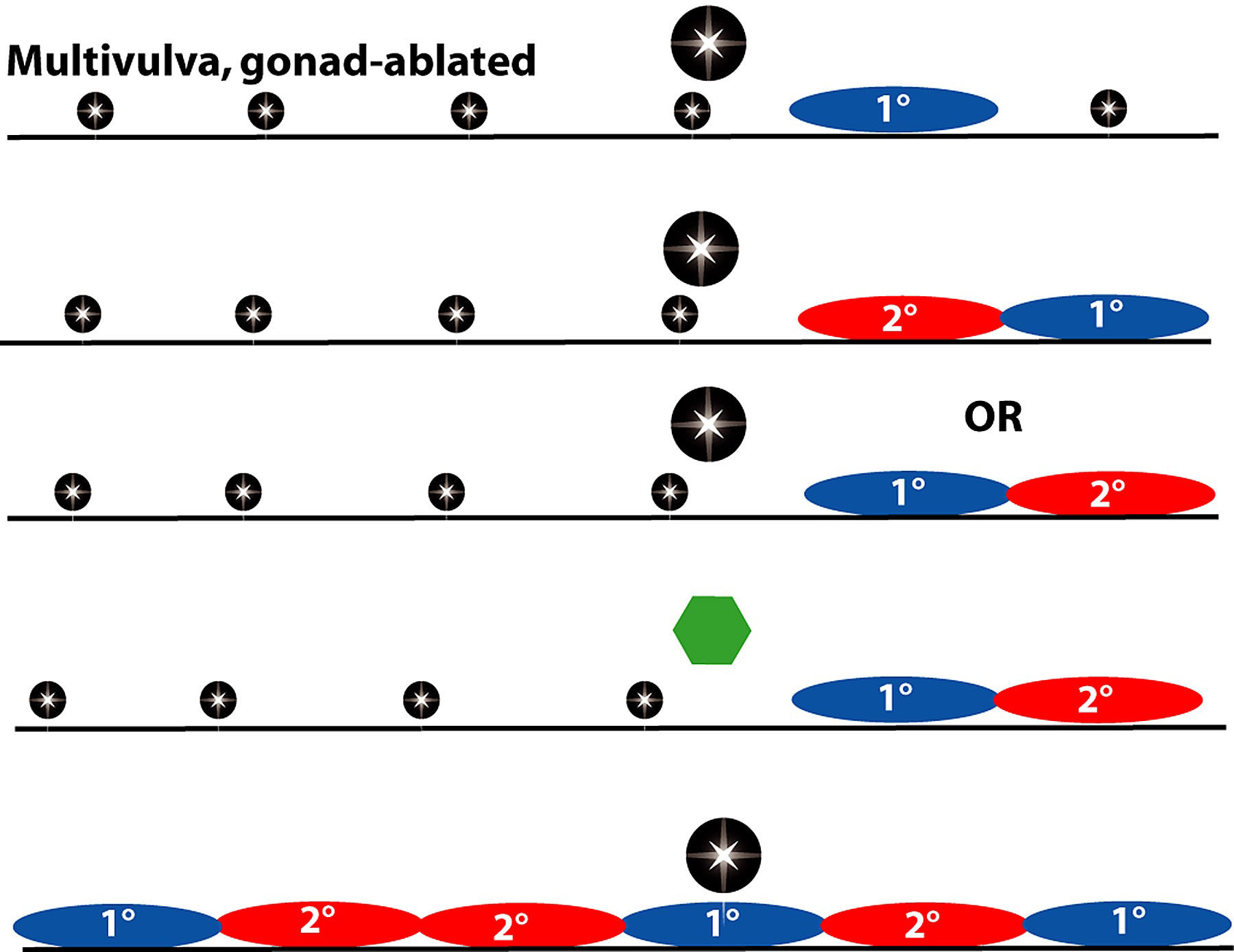
Figure 6. Pattern of VPC fates and main defects in induction. Ablation (indicated by black star) of the gonad or of the anchor cell results in a vulvaless phenotype with all six VPCs having 3° fates. Vulvaless animals have an anchor cell but vulval induction does not occur, resulting in a vulvaless phenotype. Multivulva mutants have more than three VPCs generating vulval progeny. Many have all six VPCs becoming 1° or 2° even in the absence of the anchor cell or gonad. Yellow, 3° fate; red, 2° fate; blue, 1° fate.
Ablation of all six VPCs in C. elegans did not result in P2.p or P9.p generating vulval progeny (Sulston and White, 1980; compare to Panagrellus redivivus (Sternberg and Horvitz, 1982; see Evolution of development in nematodes closely related to C. elegans), suggesting that only P3.p-P8.p form the "vulval equivalence group." Additional support for this conclusion comes from the observation that mutations that activate the LET-23 or LIN-12 signaling pathways (and hence allow VPCs to generate vulval cells independent of a signal from the gonad) only cause P3.p-P8.p but P2.p or P9.p to adopt vulval fates (Greenwald et al., 1983; Ferguson et al., 1987; Sternberg, 1988). Moreover, when the gonad position relative to the VPCs is displaced by a dig-1 mutation, the position of the 1° cell can shift, no cells other than P3.p-P8.p generate vulval cells (Thomas et al., 1990).
The hox gene lin-39 is a major determinant of the VPC group (Clark et al., 1993). lin-39 is expressed in P3.p to P8.p (Salser et al., 1993). Lack of lin-39 activity results in lack of VPCs because the presumptive VPCs fuse with the hyp7 epidermis. However, overexpression of lin-39 does not cause additional Pn.p cells such as P2.p or P9.p to become competent. lin-39 has at least two roles: preventing fusion and stimulating cell division (Figure 5). A mutation in the eff-1 results in lack of cell fusion (Mohler et al., 2002). Inactivation of both eff-1 and lin-39 results in VPCs that fail to fuse and fail to divide (Shemer and Podbilewicz, 2002). WNT signaling via BAR-1 beta-catenin is necessary to maintain expression of lin-39 in the P(3-8).p (Eisenmann et al., 1998). Expression of lin-39 is sufficient to bypass the requirement for BAR-1 function in generating competent VPCs (Eisenmann et al., 1998). Using a bar-1 mutant to sensitize the VPCs for loss of competence, Eisenmann et al. (1998) showed that let-23 and the somatic gonad were involved in establishing competence of VPCs.
sem-4, which encodes a zinc finger protein, is necessary for full lin-39 expression, and overexpression of lin-39 can rescue the defect of a sem-4 mutant (Grant et al., 2000). CEH-20 and UNC-62 are co-factors of LIN-39 and as such affect vulval development (Yang et al., 2005). Two redundant GATA factors, encoded by the adjacent egl-18 and elt-6 genes, are necessary for vulval competence, and are regulated by LIN-39 and CEH-20 (Koh et al., 2002). The translocation nT1 has a highly penetrant VPC competence defect (Ferguson and Horvitz, 1985; Ferguson et al., 1987). nT1 disrupts the egl-18/elt-6 operon, preventing its expression in the vulva (Koh et al., 2004).
More detailed analysis indicates that the VPCs, while multipotential, are not equivalent in their ability to respond to the major inductive signal (Clandinin et al., 1997). In particular P7.p and P8.p are less sensitive to EGF-R signaling than is P6.p (Figure 4). This difference in sensitivity is dependent on the hox gene mab-5, which is expressed in P7.p and P8.p but not P(3-6).p (Salser et al., 1993). Also, sem-4 is preferentially required in P7.p, and this effect is in part due to mab-5 (Grant et al., 2000). The details of fate specification may differ between P5.p and P7.p. These differences are relatively subtle, but very intriguing in light of vulval development in other nematode species (see Evolution of development in nematodes closely related to C. elegans).
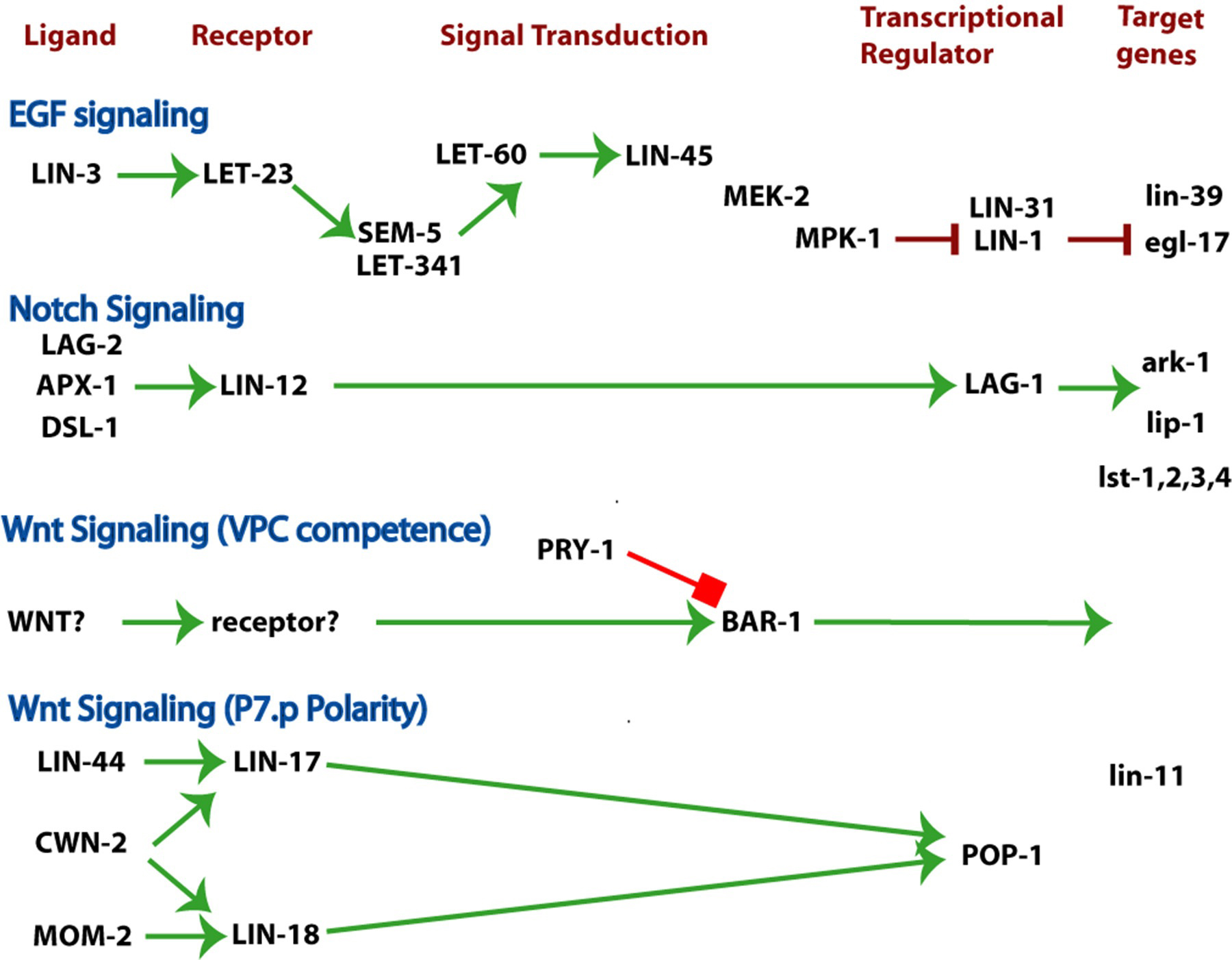
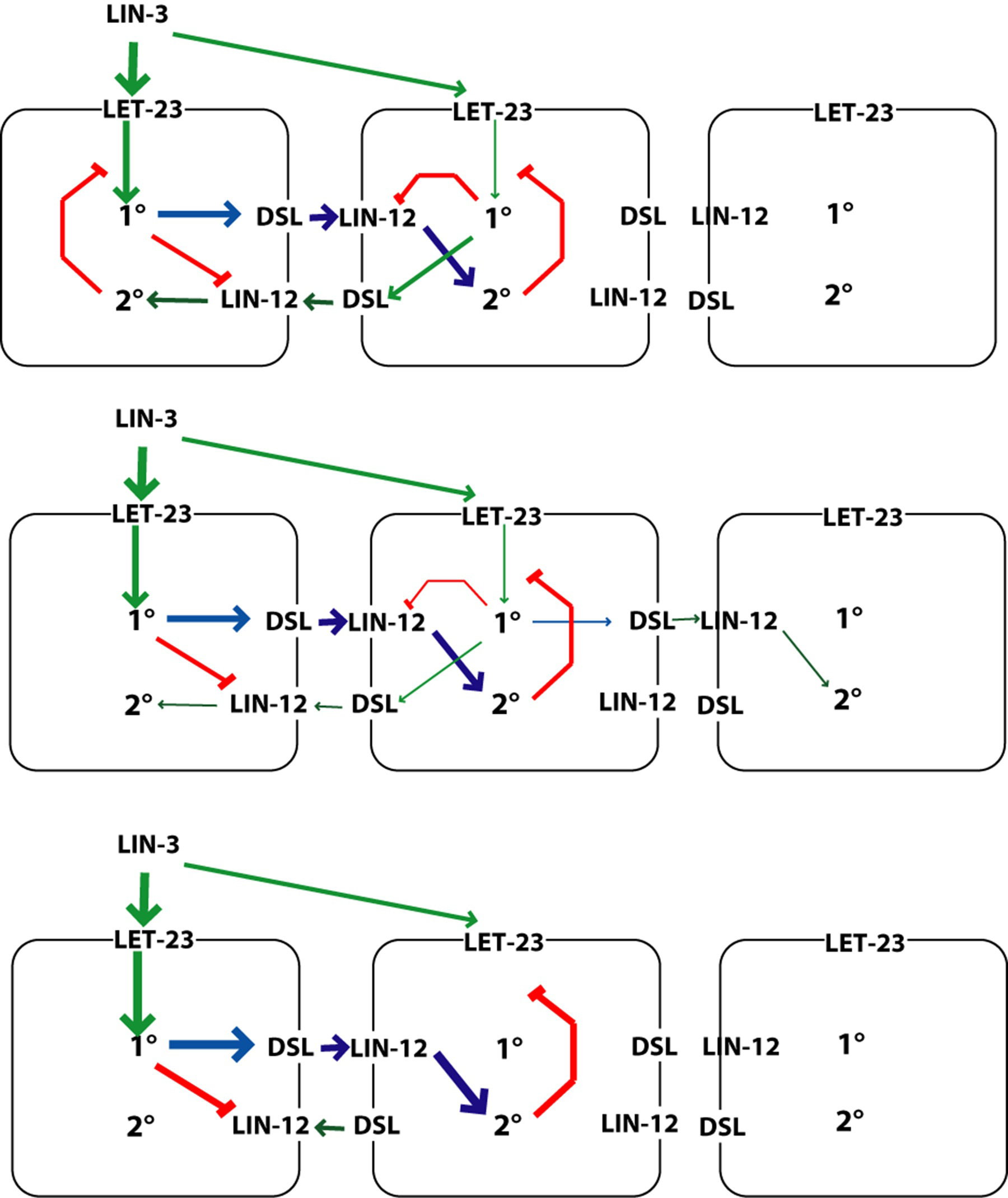
The 1°, 2°, and 3° fates occur in a precise spatial pattern. Formation of the pattern of VPC fates involves several key elements. An inductive signal from the anchor cell acts via LET-23 to induce P6.p and to a lesser extent P5.p and P7.p. Lateral signaling among induced VPCs via LIN-12 that specifies 2° fates and inhibits 1° fates. Cross inhibition exists between the two pathways: LIN-12 signaling turns on inhibitors of LET-23 signaling, and LET-23 signaling up-regulates ligand for LIN-12 and down-regulates the receptor LIN-12 (Figure 7). Inhibitors also act as if they are noise suppressors to ensure that fate specification is dependent upon signals. Figure 8 shows the key signaling pathways involved in vulval patterning. The current view of vulval precursor cell patterning is that the two systems cooperate to give a precise pattern (reviewed by Sternberg, 2004; Sundaram, 2004). Vulval induction apparently involves both a graded signal (LIN-3 acting via LET-23) as well as a sequential, subordinate signal (DSL ligands acting via LIN-12; see LIN-12/Notch signaling in C. elegans).
|
Figure 7. Three-signal model for vulval patterning. The invariant pattern of VPC fates is proposed to arise from the combined action of three signaling pathways. Yellow, 3° fate; red, 2° fate; blue, 1° fate. Green arrows, positive regulation; red bars, negative regulation. The inductive signal from the anchor cell (LIN-3; green) is necessary for non-3° fates. The Lateral signal among the induced VPCs (blue) promotes the 2° fate (via LIN-12). The negative signal is inferred from the non-autonomy of some class B synMuv genes. Adapted from Herman and Hedgecock (1990); Horvitz and Sternberg (1991).
|
Figure 8. Three major signaling pathways patterning the vulva and uterus. EGF signaling. Notch signaling. WNT signaling. The ligand, receptor(s), signal transducers, nuclear targets and regulated genes for each pathway are indicated. Wnt signaling plays several roles during vulval development, including specification of VPC competence and polarity of P7.p. See RTKRas/MAP kinase signaling, LIN/Notch signaling in C. elegans, and Wnt signaling. Green arrows, positive regulation; red bars, negative regulation.
VPC patterning is conveniently and appropriately thought of in terms of VPC level specification (Sternberg and Horvitz, 1986). A specified VPC generates a characteristic 1°, 2°or 3° lineage, and expresses markers that are characteristic of each fate.
An open question in C. elegans development concerns fate commitment. I use specification to mean a cell knows what to do but is not irreversibly committed to that fate; commitment is irreversible (within a reasonable set of perturbations). The rapidity of C. elegans development, and the lack of long-term culture have precluded extensive tests of commitment. One can argue that cells are specified and then development proceeds without any need for long-term stability. One experiment with VPCs revealed that the 2° fate VPCs are not committed (Wang and Sternberg, 1999). Expression of high levels of LIN-3 after the VPCs have divided in a LIN-12 activated mutant alters their fate such that they express 1° lineage markers and display the morphological hallmarks of 1° lineages, such invagination; thus cells specified to become 2° are not committed even after they divide. The 3° fate is irreversible after fusion with hyp7, about an hour after their cell division. The 1° fates so far have not been altered after their division (Ambros, 1999; Wang and Sternberg, 1999). Ambros (1999) found that VPCs could become specified to become 1° prior to the end of their S phase but specification of 2° fates required the end of S phase. In this view, the cell cycle helps prioritize the specification process.
Hybrid lineages arise by too little induction (for example ablation of the anchor cell before the VPCs are fully induced) or by late induction acting on the daughter cells (for example if LIN-3 expression is induced after the VPCs divide; Wang and Sternberg, 1999). Fusion of VPC daughters (P3.pa, P3.pp, P4.pa, P4.pp, P8.pa, and P8.pp) occurs about an hour after they are formed, which eliminates their competence to respond to LIN-3.
Vulva development involves cell cycle in two distinct ways. Cell cycle regulation is one important consequence of vulval cell fate specification, and conversely, the cell cycle influences fate specification. One of the major differences between the uninduced 3° and the induced vulval (1° and 2°) VPC fates is in the extent of cell division. Cyclin E, encoded by cye-1 is necessary for proliferation of the VPCs (Fay and Han, 2000). In cye-1 mutants, there are fewer VPC divisions, but differentiation, assayed by expression of egl-17::gfp occurs at the normal time. Moreover, Boxem and van den Heuvel (2002) found that cell division could be uncoupled to some extent from differentiation.
The heterochronic genes lin-4, lin-14, and lin-28 control the G1 to S transition of VPCs (Euling and Ambros, 1996). The cell cycle inhibitor cki-1 is one target of this heterochronic pathway (Hong et al., 1998). cki-1::GFP is expressed in the VPCs during their G1 phase. Inactivation of cki-1 results in VPC division during the L2 stage, and expression of cki-1 in P6.p under control of egl-17 transcriptional regulatory sequences blocks its mitosis (Hong et al., 1998). cki-1 expression in VPCs is reduced in the absence of lin-14, which results in precocious development of the VPCs.
The anchor cell (AC) induces vulval formation directly by expressing the epidermal growth factor (EGF)-like ligand LIN-3. The action of LIN-3 is graded and acts at a distance. The induction by the anchor cell can also be indirect, with induction of LIN-12-mediated lateral signaling and possibly additional lin-3 signaling.
Ablation of the anchor cell or its ancestors prior to the L3 stage completely blocks vulval development, and the VPCs have 3° fates (3°-3°-3°-3°-3°-3°). Ablation of all gonadal cells but the anchor cell results in full induction of the vulva (Kimble, 1981). Males will copulate with these vulvae but not transfer sperm (M. Barr, K. Liu and P. Sternberg, unpublished observations).
lin-3 encodes the inductive signal from the anchor cell that induces the VPCs (Hill and Sternberg, 1992; Figure 9). lin-3 is necessary for vulval induction (Horvitz and Sulston, 1980; Sulston and Horvitz, 1981; Ferguson and Horvitz, 1985; Ferguson et al., 1987; Hill and Sternberg, 1992). lin-3 is expressed in the anchor cell during the time induction takes place. Expression of the EGF domain of LIN-3 directed by a heat shock (hsp-16) promoter/enhancer is sufficient to induce the vulva in the absence of the gonad and hence the anchor cell (Hill and Sternberg, 1992).
|
Figure 9. Level of LIN-3 directs pattern of VPC cell fates. In wild-type, the six VPCs adopt the 3°-3°-2°-1°-2°-3° pattern in each case. Animals with reduced lin-3 function (lin-3 reduction of function; lin-3(rf)) display a range of phenotypes including only P6.p induced to 1°. A further decrease in lin-3 function results in a highly penetrant vulvaless phenotype. Excessive lin-3 in the anchor cell (lin-3(+ + +)) results in an expansion of the pattern, including cases as shown here. Yellow, 3° fate; red, 2° fate ; blue, 1° fate.
The anchor cell signals at a distance from the VPCs. Cell ablation studies reveal that a distal VPC can be induced to a 1° or 2° fate (or some intermediate; Sternberg and Horvitz, 1986; Katz et al., 1995; Sherwood and Sternberg, 2003). In addition, in dig-1 mutants, a dorsally located anchor cell can induce patterns such as 1°-2°-1°, 1°-2°-1°-2°or 2°-2°-2°-2° (Thomas et al., 1990). The dorsally located anchor cell can extend a process ventrally, but this occurs after induction, and is fully consistent with the anchor cell invasion of the vulval epidermis (Sherwood and Sternberg, 2003).
The anchor cell signal is able to act at a distance and appears graded based on a correlation of distance from the anchor cell and the fate of individual VPCs (Sternberg and Horvitz, 1986). An individual VPC, after ablation of other VPCs, can have a 2° fate. Also, in a dig-1 mutant there can be 2° VPCs without 1° VPCs (Thomas et al., 1990). In addition, molecular genetic experiments (Katz et al., 1995; Katz et al., 1996) strongly support the possibility of a graded signal. The dose of LIN-3 was controlled in two ways. The EGF domain of LIN-3 along with a synthetic signal sequence was expressed under the transcriptional control of a heat-shock enhancer/promoter. By varying the duration and temperature of heat shock, different apparent doses of LIN-3 could be produced. A high dose results in 1° fates; an intermediate dose in 2° fates; and a low dose in 3° fates. In most cases there is a distribution of fates as well as intermediate fates. A distinct set of experiments with partially activated LET-23 gave similar results (Katz et al., 1996).
The graded expression of egl-17, the best-known target of LET-23 signaling in the 1° VPC (Burdine et al., 1998), provides a molecular correlate for a graded inductive signal. egl-17-CFP-LacZ, is initially expressed in P5.p, P6.p and P7.p with higher expression in P6.p, thereby indicating a graded signal (Yoo et al., 2004). Its expression is increased in P6.p and decreased in P5.p and P7.p due to LIN-12 signaling as described below.
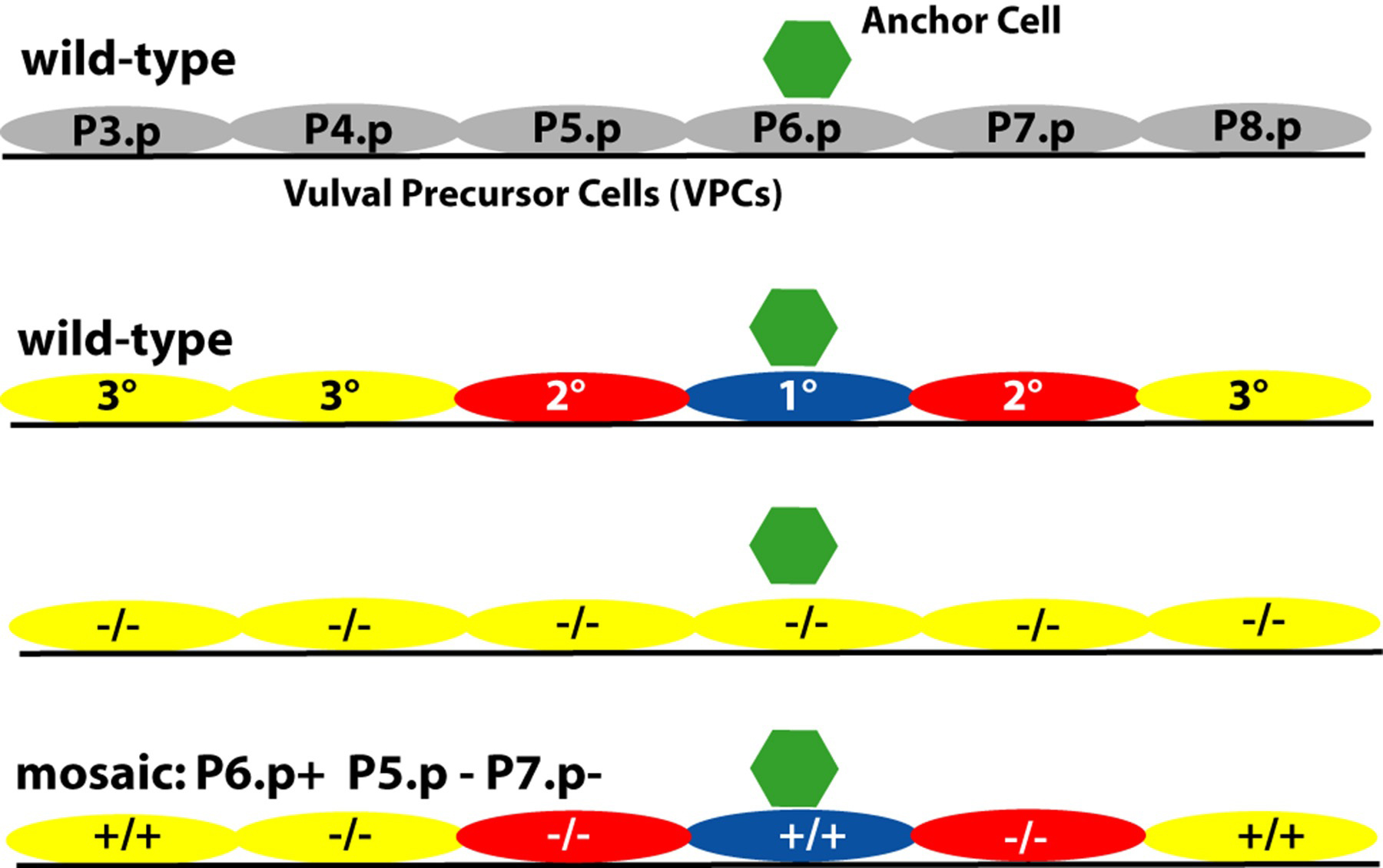
In addition to the strong support for a graded signal, there is also strong support for a sequential signaling mechanism in which activation of LET-23 in P6.p leads to activation of LIN-12 in P5.p and P7.p. Certain genetic mosaics lacking let-23 activity suggest that direct induction by the AC is not necessary for the 2° fate: if let-23(+) is present in all cells, there is a wild-type vulva; if all cells are let-23(-) there is no vulva (Figure 10; Simske and Kim, 1995; Koga et al.,1995). If P6.p is let-23(+) but P5.p and P7.p are let-23(-), then in almost all cases there was a normal vulva. Thus, a cell can become 2° without receiving LIN-3. This key result is consistent with the observations that activation of LIN-12 can specify a 2 ° fate in the absence of an anchor cell (Greenwald et al., 1983) or of LET-23 signaling (Sternberg and Horvitz, 1989; Han et al., 1990), and strongly makes the point that the direct induction of 2° by the anchor cell is not necessary. LIN-12 is expected to be the receptor for this 2°-inducing signal: lin-12 activity is necessary and sufficient for the 2° fate (Greenwald et al., 1983), and ligands for LIN-12 are up-regulated in P6.p in response to the inductive signal (Chen and Greenwald, 2004).
|
Figure 10. Sequential signaling inferred from mosaic analysis of let-23. yellow, 3° fate; red, 2° fate; blue, 1° fate. Genotype of VPCs in key mosaic animal shown. -/-, lacking let-23 activity; +/+, let-23(+) activity present. In the absence of let-23, all VPCs become 3°. In mosaic animals which lack let-23 in P5.p and P7.p but have let-23 in P6.p, the vulva is usually wild-type. This observation indicates that let-23(+) is not required in presumptive 2° VPCs. Results of Simske and Kim (1995) and Koga and Ohshima (1995).
The converse mosaic animals had a striking phenotype: if P6.p is let-23(-) but P5.p and P7.p are let-23(+), then additional cells are induced (Simske and Kim, 1995; also see Hajnal et al., 1997). Hajnal et al. argue that LET-23 might be sequestering LIN-3 and preventing its spread to more distal cells.
Dutt et al. (2004) have suggested that lin-3 expression in induced VPCs extends the range of induction, and that this role of lin-3 depends on ROM-1. rom-1 encodes a protease related to Drosophila Rhomboid, which cleaves Spitz, a Drosophila protein related to LIN-3. lin-3 encodes multiple isoforms that differ in the region between the EGF domain and the transmembrane domain (Hill and Sternberg, 1992; Dutt et al., 2004), a part of the protein that would likely be cleaved for LIN-3 in a relay mechanism. The B but not the A isoform of LIN-3 is ROM-1 dependent (Dutt et al., 2004).
Two transcription factors, LIN-31 and LIN-1 are likely targets of the MAPK pathway (Tan et al., 1998). LIN-31 and LIN-1 can both be phosphorylated by MAP kinase MPK-1. Inactivation of either lin-1 or lin-31 leads to a Multivulva phenotype. LIN-1 and LIN-31 form a complex that is disrupted by LIN-31 phosphorylation (Tan et al., 1998). Increased expression of LIN-31 or a non-phosphorylatable LIN-31 protein causes a vulvaless phenotype. During the L3 stage, lin-31 mutants display both vulvaless and multivulva phenotypes (Ferguson and Horvitz, 1985). lin-31 mutants display inappropriate divisions of Pn.p cells during the L2 stage (Ferguson et al., 1987; Miller et al., 2000), suggesting also a role in the proper specification of VPCs.
lin-39 is upregulated by LET-23 signaling (Maloof and Kenyon, 1998) and is an excellent candidate for a direct target. Since overexpression of lin-39 does not result in gonad-independent vulval differentiation, it is unlikely to be the only target (Figure 5). egl-17 is induced early in the induction process (Burdine et al., 1998; Ambros, 1999; Wang and Sternberg, 1999; Cui and Han, 2003; Yoo et al., 2004) and could be an early target of RAS-stimulated transcription. Romagnolo et al. (2002) used RNA from whole animals hybridized to microarrays to search for RAS-regulated genes and found 718; among these might be some that are physiologically relevant during vulval development.
Mediator complex proteins, which link site-specific transcription factors to RNA polymerase, also are necessary for vulval development. lin-25 and sur-2 (mdt-23) are defective in vulval induction and also have defects earlier and later during vulval development (Ferguson et al., 1987; Tuck and Greenwald, 1995; Singh and Han,1995). mdt-6 is positive-acting (Kwon and Lee, 2001). It is striking that other Mediator components have negative effects, e.g., dpy-22 (mdt-12; Moghal et al., 2003; H. Sawa, unpublished observations). There are two extreme interpretations of this observation. First, there might be differential requirements for mediator components on distinct transcriptional targets. Second, there might be one main target for Mediator and there is a regulatory pathway acting among the Mediator components with some components acting in a positive and others in negative manner.
In addition to its role in VPC competence, activation of WNT signaling can increase the extent of vulval induction if the LET-23 pathway is compromised (Gleason et al., 2002). It is not known whether WNT signaling can compensate for the complete absence of LIN-3 signaling.
The invariance of vulval development is not only due to the growth of C. elegans under standardized laboratory conditions; however, when vulval development is partly compromised, a number of physiological effects are evident. Ferguson and Horvitz (1985) observed that certain vulvaless mutations are suppressed by starvation during larval growth while others are suppressed by passage through the dauer phase. The suppression does not appear to correlate with genetic pathways, and might be due to some type of informational suppression (see Genetic suppression). Recently several other potentially physiological inputs (Battu et al., 2003; Moghal et al., 2003), and an important role for zinc have been identified (Bruinsma et al., 2002; Yoder et al., 2004).
The heterotrimeric G protein Gq (EGL-30; see Heterotrimeric G-proteins in C. elegans) stimulates vulval induction under certain growth conditions, in particular growth in liquid (Moghal et al., 2003). Expression of activated EGL-30 in neurons stimulates vulval induction. The EGL-30 effect requires the voltage-gated calcium channel encoded by egl-19. The site of action of egl-19 for this effect is in muscle cells. These observations lead to the hypothesis that neuronal and muscle excitation can somehow affect vulval development. The positive signal from Gq acts in parallel to or downstream of RAS. Because the effect is of the magnitude of WNT signaling via BAR-1 β-catenin, it is possible that this is effect is via the WNT pathway (Moghal et al., 2003).
A G protein coupled receptor, SRA-13, acts negatively on vulval development via the G protein GPA-5 (Battu et al., 2003). Starvation decreases vulval induction, and this effect depends upon SRA-13. sra-13 is expressed in chemosensory neurons as well as body-wall muscle (Battu et al., 2003) and gpa-5 is also expressed in chemosensory neurons. The sites of action of SRA-13 and GPA-5 for vulval induction effects are not known. The relationship of the positive EGL-30 (Gq) effect and negative GPA-5 effects is intriguing but not yet known. These findings highlight the importance of nematode physiology, revealed by growth conditions, on vulval development. The effect on vulval induction of growth in liquid (Moghal et al., 2003) and of starvation (Battu et al., 2003) were only revealed by using sensitized genetic backgrounds. I believe that there could be much plasticity lurking beneath the surface, a surface that can be scratched away by careful genetic analysis.
Two genes identified by loss-of-function mutations that suppress activated LET-60 Ras encode cation diffusion facilitators that appear to regulate zinc. CDF-1 is a cation diffusion regulator, and has sites of action in the intestine as well as in the VPCs, consistent with it expression in both tissues (Bruinsma et al., 2002). Exogenous zinc negatively regulates LET-23 signaling (Bruinsma et al., 2002). SUR-7 encodes a distinct cation diffusion facilitator (Yoder et al., 2004). Yoder et al. (Yoder et al., 2004) argue that zinc regulation is at the level of KSR-1 and PAR-1, which somehow regulate LIN-45 Raf signaling (see RTKRas/MAP kinase signaling).
A variety of negative regulators of vulval induction have been identified. Their roles in signal transduction by LET-23 are described in RTKRas/MAP kinase signaling.
The synMuv genes are three classes (A, B and C) of broadly expressed nuclear proteins that in various combinations inhibit VPCs from adopting vulval fates. Ferguson and Horvitz (1989) described two redundant sets of genes, class A and B, where hermaphrodites defective in both A and B function have a Multivulva phenotype. Mutants defective in only A, or in only B, have normal vulval development. Molecular cloning revealed that many Class B synMuv genes encode proteins with mammalian orthologs that have been well-characterized as transcriptional regulators, including the synMuv genes lin-35 Rb , efl-1 E2F, and hda-1 histone deacytelase (Lu and Horvitz, 1998), as well as a zinc finger protein (Melendez and Greenwald, 2000) and components of the NuRD and SWI/SNF complexes (Lu and Horvitz, 1998; Solari and Ahringer, 2000; Ceol and Horvitz, 2001; Cui et al., 2004). Other synMuv genes encode novel nuclear proteins.
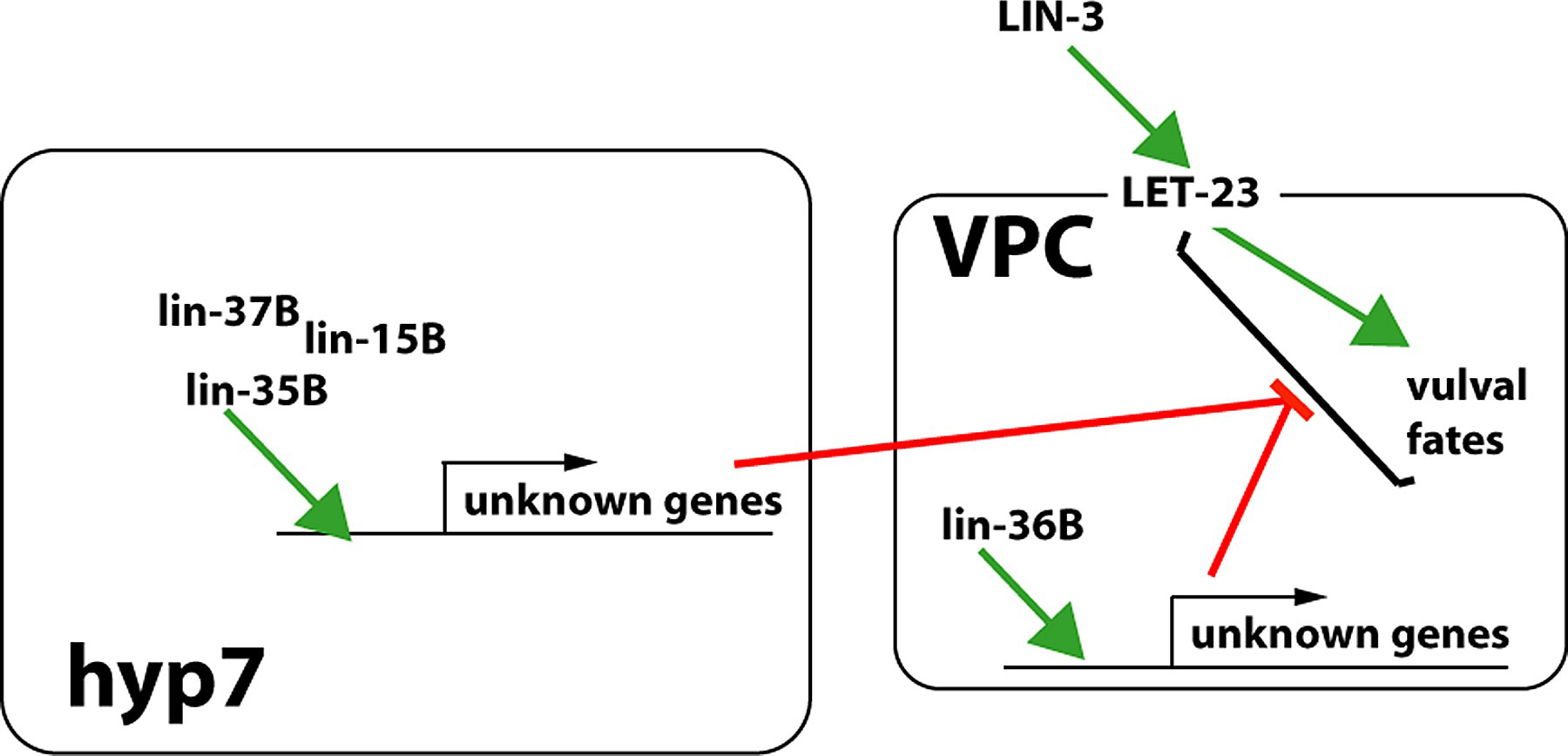
Epistasis experiments suggest that synMuv genes inhibit LET-23 pathway output (Ferguson et al., 1987; Huang et al., 1994). Site of action studies with synMuv genes suggest that they control a pathway of intercellular signaling from hyp7 to the VPCs, although no signal has been identified (Figure 11; Figure 12). lin-15AB and lin-37B mosaics suggested action not in the VPCs, and possibly in hyp7 (Herman and Hedgecock, 1990; Herman and Hedgecock, 1995); Mosaic analysis and tissue-specific rescue experiments indicate that the focus of lin-35 is hyp7 (Myers and Greenwald, 2005). However, lin-36B mosaics suggest a site of action in the VPCs (Thomas and Horvitz, 1999), and lin-15A mosaics and tissue-specific rescue experiments also suggest a VPC focus (A. Gonzalez-Serricchio and P. W. Sternberg, unpublished observations).
|
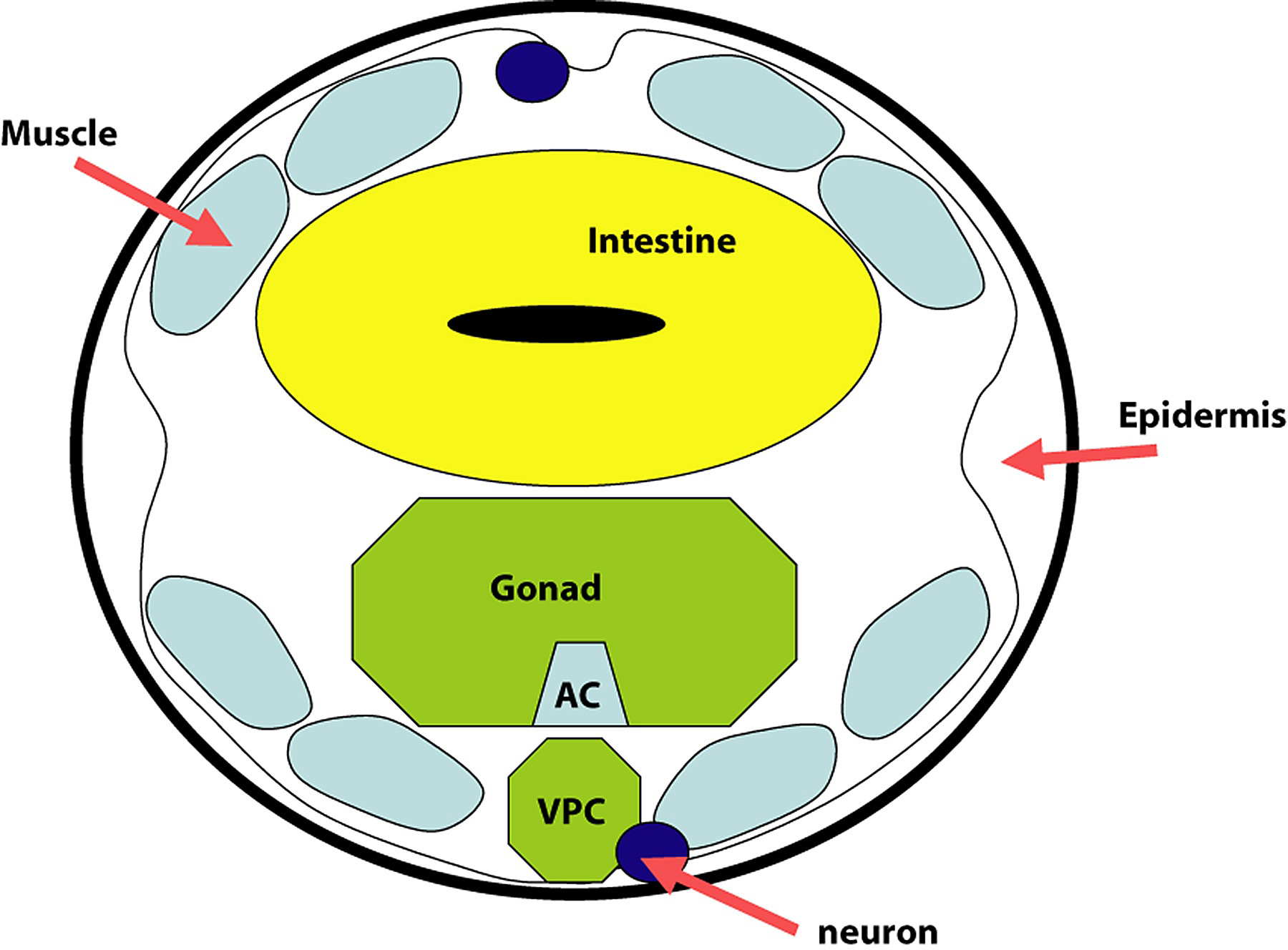
Figure 11. Schematic transverse section through the developing vulva. The major tissues of the worm at the position of the vulva are shown in this schematic transverse section. VPC, vulval precursor cell (P6.p); AC, anchor cell. The VPCs are separated from the AC by the basement membranes of both the gonad and the epidermis, and the pseudocoelom (body cavity). See WormAtlas for detailed description of the anatomy.
|
Figure 12. Synthetic multivulva mutants suggest signaling from hyp7 to VPCs. lin-15b, lin-35 and lin-37 have been argued to act in hyp7 while lin-36 has been argued to act in the VPCs. These proteins where known are nuclear and presumably regulate gene expression (green arrows), although the physiological targets are unknown (unknown genes) as is the sign of the regulation. Epistasis analysis suggests that the synMuv genes somehow act negatively on LET-23 signaling (red bars) thereby inhibiting the specification of 1° and 2° VPC fates.
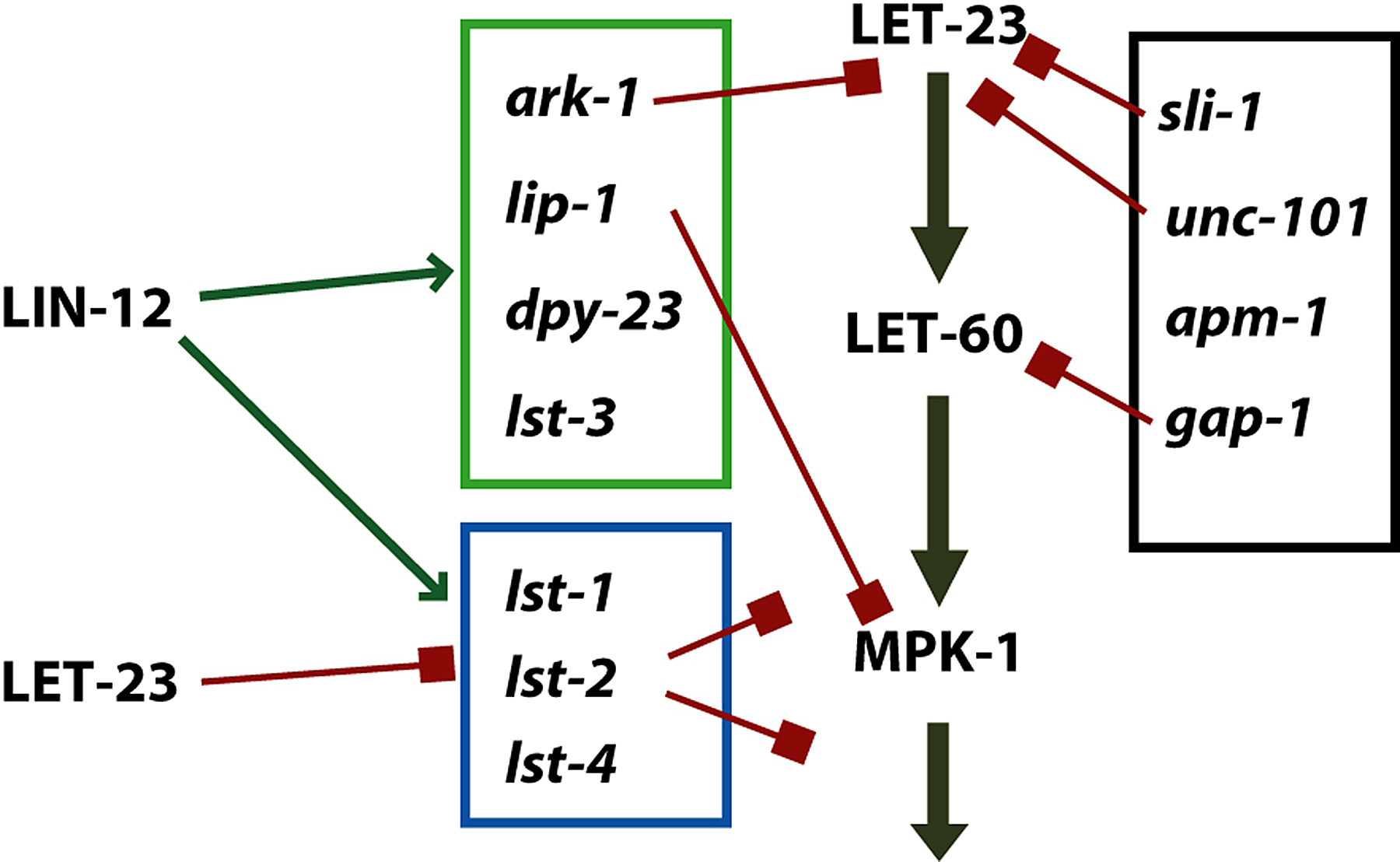
Other negative regulators were identified as suppressors of hypomorphic mutations in LET-23-RAS pathway components. These genes do not interact strongly with the synMuv genes, but do interact with each other. sli-1, unc-101, ark-1, gap-1 and lip-1 (Jongeward et al., 1995; Lee et al., 1994; Yoon et al., 1995; Hajnal et al., 1997; Berset et al., 2001; Hopper et al., 2000) are of this class. In addition, Yoo et al. (2004) found four additional negative regulators, dpy-23, lst-1, lst-2, lst-3, and lst-4. These four regulators, and lip-1 (Berset et al., 2001) are targets of LIN-12 lateral signaling.
unc-101 and apm-1 each encode medium chains of the AP-1 adaptin, and these are inhibitory on vulval induction (Lee et al.,1994; Shim, 2000); while LET-23 is a plausible target for these membrane trafficking proteins, the targets are not known. dpy-23 encodes a medium chain of the AP-2 adaptin and is induced by LIN-12 (Yoo et al., 2004; see below).
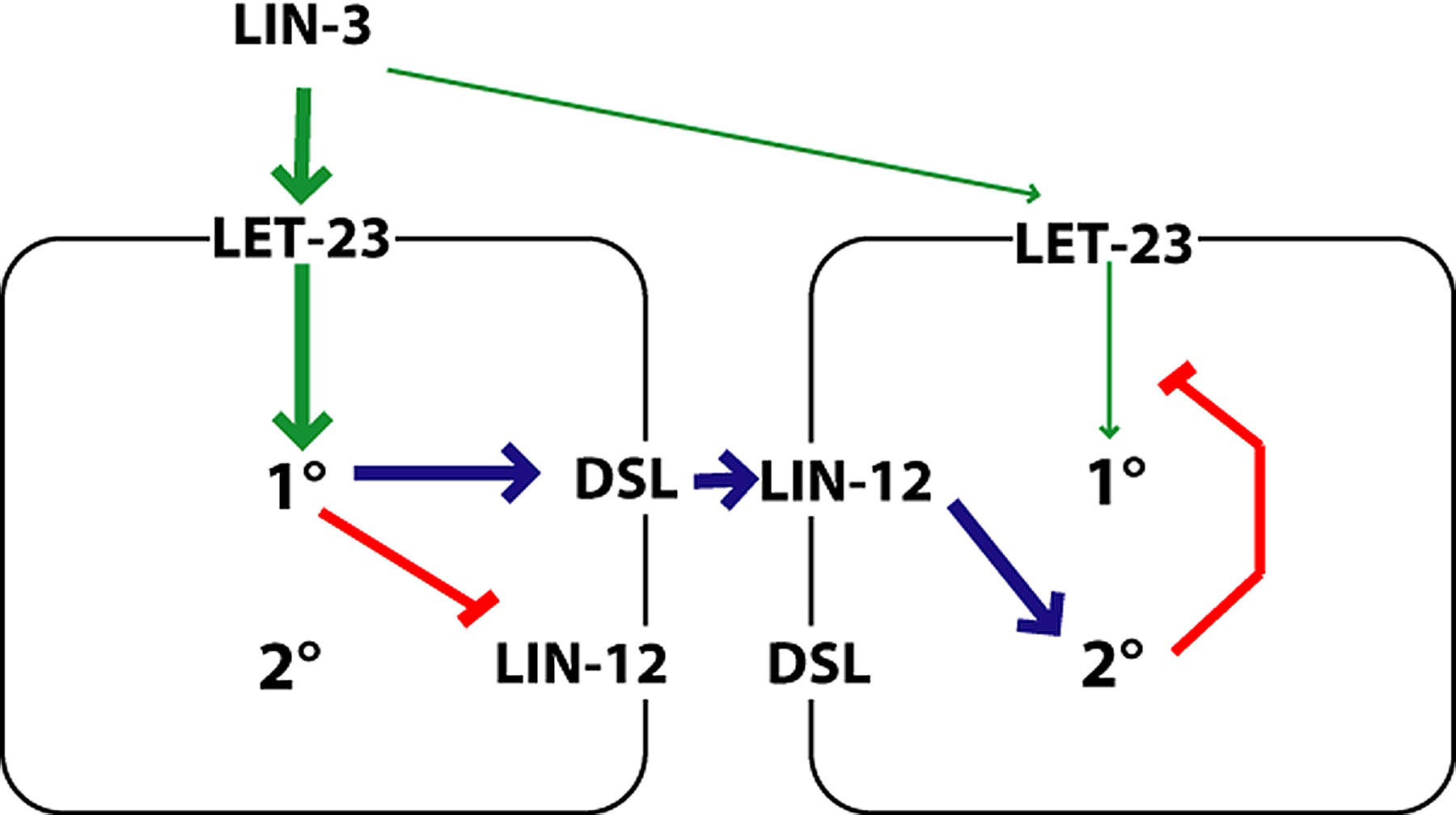
The VPCs signal each other via a LIN-12-mediated lateral signaling pathway. This pathway has two related roles: to prevent adjacent induced VPCs from both becoming 1°, and to induce VPCs that flank a 1° VPC to become 2° (Figure 13).
|
Figure 13. Simple model of antagonism between 1° and 2° VPCs. Signals for 1° specification (green) compete with signals for 2° specification (blue). In this example, the left cell is winning the competition to become 1° because it is receives more LIN-3. LIN-3 signaling via LET-23 promotes the 1° fate, which includes the production of DSL ligands and downregulation of the receptor LIN-12. DSL ligands activate LIN-12 on neighboring cells, thereby promoting the 2° fate, which inhibits LET-23 signaling. Red bars, inhibition. Model based on Sternberg and Horvitz (1989); Berset et al. (2001); Shaye and Greenwald (2002); and Yoo et al. (2004).
lin-12 is necessary and sufficient for specification of the 2° VPC fate (Greenwald et al., 1983). Null alleles of lin-12 lack 2° VPCs. Gain of function lin-12 mutants cause all six VPCs to generate 2° lineages, even in the absence of inductive signaling. As described in LIN-12/Notch signaling in C. elegans, lin-12 encodes a Notch-type transmembrane receptor (Yochem et al., 1988).
lin-12(lf) hermaphrodites have additional anchor cells, and this complicated somewhat the analysis of vulval induction (Greenwald et al., 1983). With two (or more) anchor cells, P5.p, P6.p and P7.p are 1° and P4.p and P8.p occasionally are 1° or one daughter is 1° like (Sternberg and Horvitz, 1989).
The lateral signal involves at least three redundant genes, apx-1, lag-2 and dsl-1 (Chen and Greenwald, 2004; see LIN-12/Notch signaling in C. elegans). APX-1 and LAG-2 are transmembrane ligands for LIN-12. DSL-1 is a secreted ligand for LIN-12. Expression of lag-2 occurs in all VPCs prior to induction and is upregulated in P6.p. Expression of apx-1 and dsl-1 appears to be initiated by induction of P6.p. The main effect is to induce neighbors to become 2°, thereby inhibiting them from becoming 1°. The redundancy of lag-2, dsl-1 and apx-1 for VPC patterning appears to have prevented the discovery of their role by genetic screens. The targets of LIN-12 include a number of redundant negative regulators of LET-23 signaling (Yoo et al., 2004; Figure 14).
|
Figure 14. Lateral signaling targets and negative regulation of LET-23 signaling. Green box, targets whose expression is induced by LIN-12; blue box, targets whose expression is inhibited by LET-23 but induced by LIN-12; Black box, targets whose expression has not been analyzed or that are not regulated by these signaling pathways. See Yoo et al. (2004) for details.
The pattern of VPC fates in a lin-15 multitvulva mutant (1°-2°-2°-1°-2°-1°; Figure 15) is reminiscent of spacing patterns such as that of bristles in the insect abdomen and thus suggests a lateral inhibition mechanism whereby a specialized cell type (1° VPC) inhibits adjacent cells from also becoming of that type and instead differentiating as some alternative (2° VPCs). While adjacent 2° VPCs are common, adjacent 1° VPCs are rare. In a lin-15 strain, which appears to have activat ed LET-23 signaling, a single VPC (e.g., P7.p) will become 1° in the absence of the gonad and of its neighbors (Sternberg, 1988). If there are two adjacent VPCs (P7.p and P8.p), they will become 1°-2°or 2°-1°, with approximately equal probability. Thus, these cells signal each other.
In the presence of the gonad, P7.p becomes 1° and P8.p becomes 2°, indicating that the inductive signal can bias the stochastic lateral signaling among induced VPCs (Sternberg, 1988). Double mutant combinations among inductive signaling pathway genes and lin-12 mutations support this view: a lin-12 loss-of-function mutation eliminates the lateral inhibition in a lin-15 mutant resulting in all 1° VPCs (Sternberg and Horvitz, 1989).
|
Figure 15. Lateral signaling. In the absence of the gonad and other VPCs, P7.p becomes 1° in a lin-15 multivulva mutant. If P8.p is also present, the two cells adopt a 1°-2° or 2°-1° pattern at random. In the presence of the anchor cell, P7.p becomes 1° and P8.p 2° (Sternberg, 1988).
The antagonism of LET-23 and LIN-12 is fundamental to the specification of 1° and 2° fates (Figure 16). As described above, LET-23 activation leads to 1° fates at high levels and 2° at low levels. The genetic requirements for this 2° specification has not been elucidated. LIN-12 lateral signaling is able to prevent adjacent cells from becoming 1° (Sternberg, 1988), and can induce a cell to become 2° (Greenwald et al., 1983; Simske and Kim, 1995; Koga and Ohshima, 1995). A presumptive 1° cell downregulates LIN-12 protein (Shaye and Greenwald, 2002). Conversely, a presumptive 2° cell downregulates LET-23 signaling (Berset et al., 2001; Yoo et al., 2004). C. elegans vulva development was the first case where the interplay of growth factor receptor and Notch signaling integration was discovered.
Activation of LIN-12 bypasses the requirement for the gonad or for LET-23 (Greenwald et al., 1983; Sternberg and Horvitz, 1989). However, a strong LIN-3 signal can override the effect of activated LIN-12, if a VPC is near the anchor cell (Sternberg and Horvitz, 1989), if lin-1 is removed (Beitel et al., 1995), or if LIN-3(EGF) is delivered by a heat shock construct (Wang and Sternberg, 1999). Validated targets of LIN-12 in presumptive 2° cells are lip-1, encoding MAPK phosphatase (Berset et al., 2001), dpy-23 and the lst genes (Yoo et al., 2004). lst-1, lst-2 and lst-4 also appear to be inhibited by LET-23 (Figure 14), which creates the potential for interesting dynamics in the cross-t alk between LIN-12 and LET-23 signaling pathways.
C. elegans VPC patterning involves multiple sources of information each of which would be capable, in principle, of generating a normal pattern. These include: the graded action of lin-3 from the anchor cell, sequential signaling from P6.p via LIN-12, sequential ROM-1-dependent LIN-3 signaling; and non-equivalence of the VPCs due to HOX gene action. The fact that WNT signaling can increase vulval induction (Gleason et al., 2002) and that WNTs are expressed in the anchor cell (Inoue et al., 2004) raises the possibility that WNT signaling is another way in which the vulval might be induced. These pathways might act in combination to generate an exquisitely precise pattern. In C. elegans the graded anchor cell LIN-3 signal and LIN-12-mediated lateral signaling are the major players. In other species, the other pathways might predominate.
|
Figure 16. Model of VPC pattern formation. Three of the six VPCs (P6.p, P7.p and P8.p) are shown in this simplified diagram to illustrate the critical aspects of the 1°-2° patterning mechanism. LIN-3 from the anchor cell acts in a graded fashion with P6.p receiving more signal than P7.p. LET-23 activation promotes the 1° fate and production of DSL ligands for LIN-12, and inhibits LIN-12 protein levels. LIN-12 activation promotes the 2° fate and inhibits the response to LET-23 activation. The three panels give a flavor of the type of dynamics that might underlie this process. Model from Sternberg and Horvitz (1989); Berset et al. (2001); Shaye and Greenwald (2002); Yoo et al. (2004) and Sternberg (2004).
There are seven vulval cell types formed from the fusion of defined cells produced by vulval lineages: vulA, vulB1, vulB2, vulC, vulD, vulE and vulF. They each have distinct functions and patterns of gene expression (Burdine et al., 1998; Sharma-Kishore et al., 1999; Inoue et al., 2002; Kiroauc and Sternberg, 2003; Inoue et al., 2005). For example, the vulval muscles vm1 insert between the vulE and vulF cells, while the vm2 muscles insert between the vulC and vulD cells. The vulF cells are the main contact with the anchor cell and then uv1 (Newman et al., 1996; Sherwood and Sternberg, 2003).
Transcription factors encoded by egl-38 (Chamberlin et al., 1997), cog-1 (Palmer et al., 2002), and lin-11 (Freyd et al., 1990) are responsible in part for the identities of specific vulval cell types. egl-38 is necessary for expression of lin-3 in vulF cells (Chang et al., 1999), and is involved in repressing egl-17 and zmp-1 in these same cells (Inoue et al., 2005). cog-1, which is expressed in all the terminal vulval cells but with different levels and dynamics, regulates gene expression in vulB1, vulB2, vulC, vulD and vulE (Palmer et al., 2002; Inoue et al., 2005). lin-11, which is more highly expressed in the vulC and vulD cells than in vulA and vulB cells, regulates gene expression in mature vulval cell types (Freyd et al., 1990; Gupta et al., 2003). The transcription factor LIN-29 has been proposed to regulate vulval cell-type genes in a temporal window (Newman et al., 2000; Inoue et al., 2002; Inoue et al., 2005).
The 1° lineage generates two cell types, vulE and vulF, in a mirror symmetric pattern EFFE around the anchor cell (Figure 17). The anchor cell is necessary for this pattern after it induces the 1° VPC (Wang and Sternberg, 2000). WNT receptor LIN-17 is necessary as well, but the phenotype of a lin-17 mutation has only been seen in the absence of the anchor cell. Transcription factor EGL-38 is necessary for lin-3 expression in the vulF cells (Chang et al., 1999), which signals to the uterine uv1 cells (Figure 18). The peptidase-like protein EGL-26 is expressed in the membranes of vulE cells and is necessary for vulF morphogenesis (Hanna-Rose and Han, 2002). Transcription factor COG-1, expressed in most vulva cells, has a related phenotype to EGL-26 (Palmer et al., 2002), but the relationship among these genes is not known.
|
Figure 17. Patterning of the 1° lineage. A. Model posits a signal via RAS to specify vulF cells near the anchor cell. B. Intact, wild-type 1° lineage has two F cells near the anchor cell and two E cells more distal. C. After AC ablation, the patterns are variable. D. In a lin-12(lf) mutant in which all ventral uterine cells but one anchor cell have been ablated, and in which P7.p has been ablated to keep only two 1° lineages, there is a normal pattern at the anchor cells, but a variable pattern more distal. Data and model summarized from Wang and Sternberg (2000).
|
Figure 18. uv1 specification. The vulF cell expresses LIN-3 and signals the presumptive uv1 cells via LET-23 and LET-60. The site of EGL-38 action is not known, but it is necessary for lin-3 expression in the vulF cell (Chang et al., 1999). Activated LET-23 bypasses this requirement for EGL-38 function.
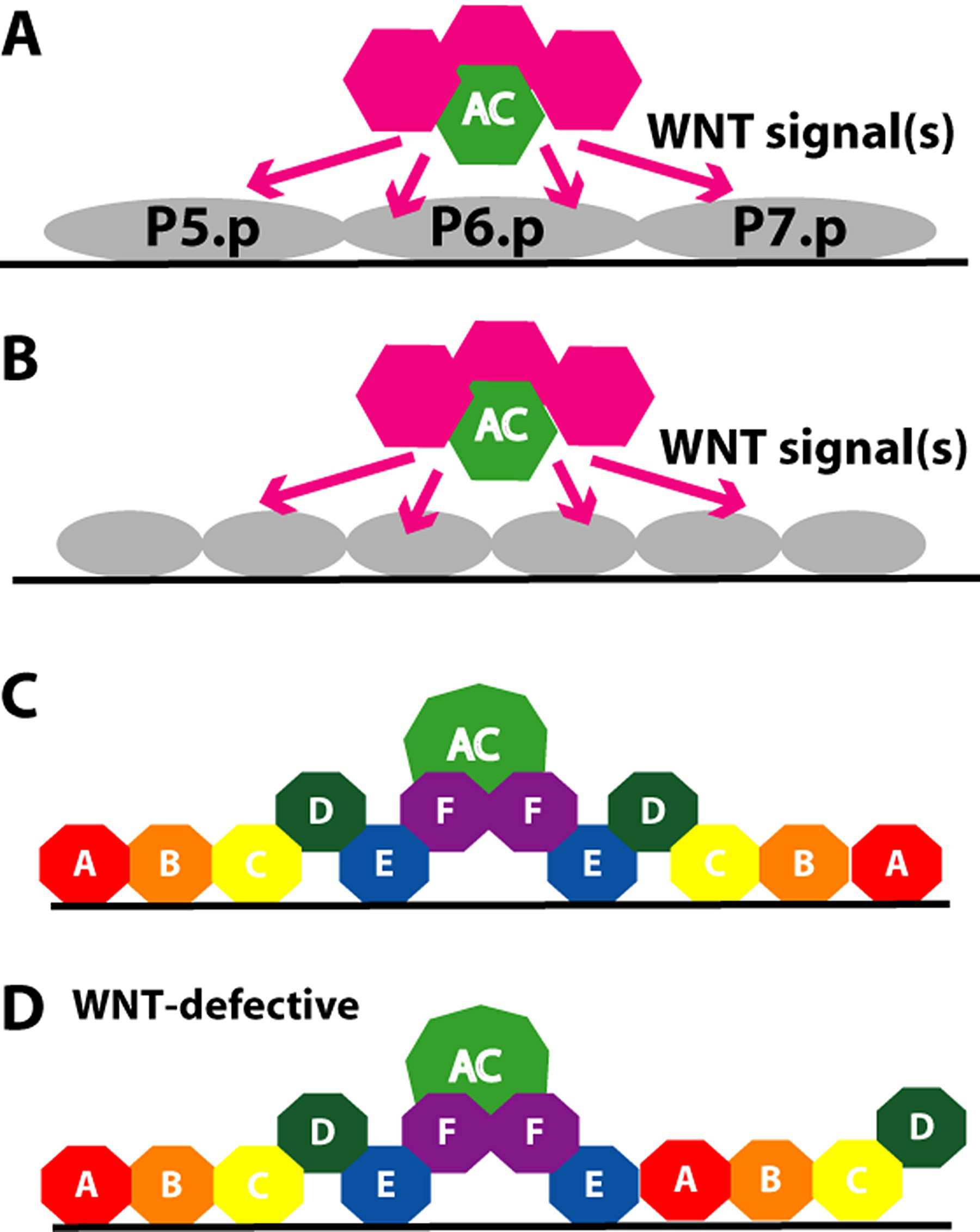
The cell division pattern and cell fates of the P5.p and P7.p lineages are mirror images, ABCD and DCBA respectively (Figure 19). The asymmetry within in each of these lineages is independent of the gonad, as for example, a single VPC in a gonad-ablated let-23(gf) mutant has asymmetry (Katz et al., 1996). WNT signaling, likely from the gonad, reverses polarity of P7.p (Inoue et al., 2004). The WNT protein MOM-2 is expressed in the anchor cell (Inoue et al., 2004) and LIN-44 might be expressed in the anchor cell or uterus based on weak GFP signal (Inoue et al., 2004) and in situ hybridization (Y. Kohara, personal communication). There are two parallel genetic pathways: one involves the WNT protein LIN-44 and the Frizzled type receptor LIN-17; the second involves the WNT MOM-2 and the Ryk type receptor LIN-18 (Ferguson et al., 1987; Sternberg and Horvitz, 1988; Sawa et al., 1996; Inoue et al., 2004). The levels of TCF/LEF protein POP-1, a WNT target, correlates with WNT signaling: P5.p daughters express POP-1 in a High-Low (HL) pattern, and P5.p granddaughters express in an HLHL pattern, while P7.p has a LH then a LHLH pattern of protein level (Deshpande et al., 2005). Thus, the pattern of POP-1 expression in the P7.p neage is reversed from the default by WNT signaling through LIN-17 and LIN-18.
|
Figure 19. Polarity of P7.p 2° lineage. Multiple WNT signals from the anchor cell and probably uterus reverse the polarity of the P7.p lineage such that it has an DCBA pattern rather than the default ABCD pattern as does P5.p. A. early L3. B. mid-L3. C. late L3. D. late L3 in animals defective in lin-17 and lin-18 WNT-receptor pathways. Model from Inoue et al. (2004) and Deshpande et al. (2005).
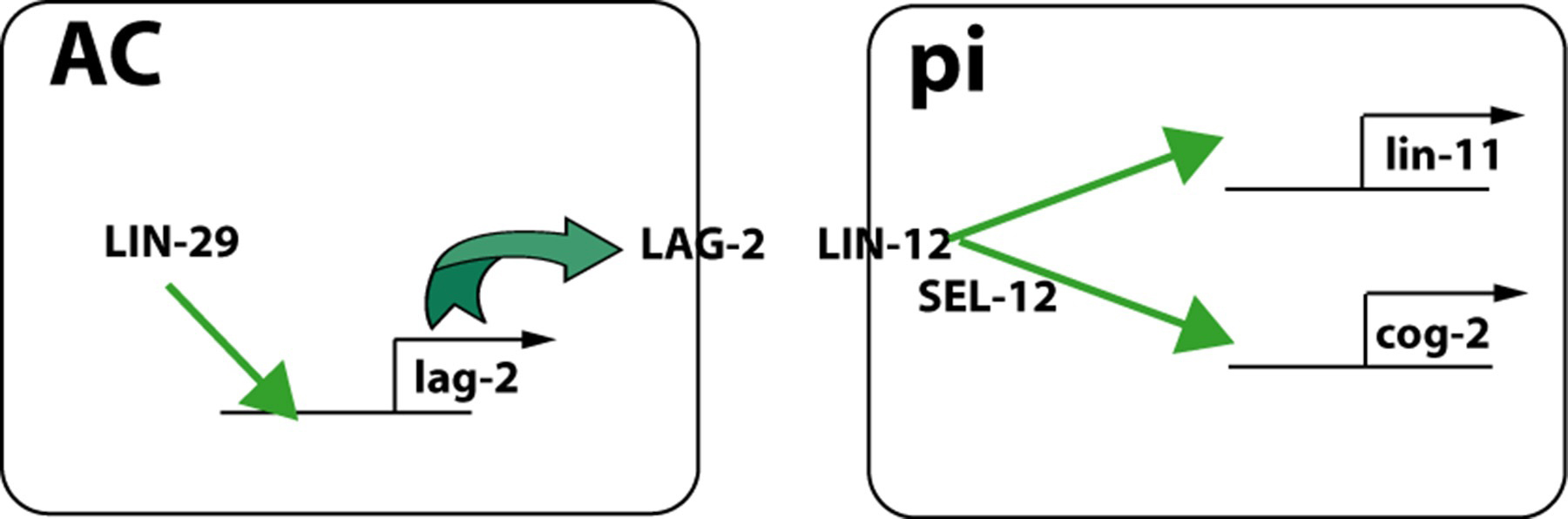
The anchor cell is the key organizer of the vulva and the connection to the somatic gonad (Newman et al., 1996; Figure 1). Once specified during the L2 molt (see Hermaphrodite cell-fate specification), the anchor cell induces and patterns the vulva via LIN-3 and LET-23. The anchor cell then patterns the ventral uterus, inducing the uterine pi cells via LAG-2 and LIN-12 signaling (Newman et al., 1995; Cinar et al., 2003; Figure 20).
|
Figure 20. Uterine pi cell induction. The anchor cell expresses DSL-type ligand LAG-2 in a LIN-29-dependent manner and signals presumptive pi cells via LIN-12. LIN-12 activation leads to expression of lin-11 and cog-2, two transcription factors necessary for utse development, including fusion of the anchor cell with utse. Model adapted from Newman et al. (1999).
egl-13 (a.k.a. cog-2), which encodes Sox domain transcription factor expressed in the uterine pi cells, is necessary for the fusion of the anchor cell and the utse (Hanna-Rose and Han, 1999; Cinar et al., 2003). lin-11, another transcription factor, is also required for differentiation of pi cells progeny and is regulated by LIN-12 signaling in pi cells (Newman et al., 1999). Sumoylation of LIN-11 is necessary for its function in pi cell specification (Broday et al., 2004).
The anchor cell induces six of 12 VU granddaughters to become pi cells, which are distinguished from the uninduced VU.xx cells by the lineage they generate. The pi cell undergo only one round of cell divisions while the uninduced cells undergo two rounds of cell division. The pi cells generate the four uv1 cells that connect to the vulF cells. Specification of the uv1 cells requires vulF cells, which express lin-3 (Chang et al., 1999). The anchor cell apparently responds to cues from vulF (and possibly negative cues from vulE) to locate the vulF cells (Sherwood and Sternberg, 2003). Meanwhile, the anchor cell helps pattern the vulE and vulF cells via LET-60 RAS (Wang and Sternberg, 2000). Moreover, a WNT signal, likely from the anchor cell reverses the polarity of the P7.p lineage (Inoue et al., 2004; Deshpande et al., 2005). The anchor cell then fuses with the utse syncytium (Newman et al., 1996). The relative timing of the signaling involved in these events has not been fully resolved.
The anchor cell breaks down the basement membranes, secreted by the gonad and the epidermis, that separates these two tissues. The anchor cell then extends a process towards the developing 1° lineage. This invasion requires the vulva; although in ~20% of vulvaless animals, the anchor cell will nonetheless invade the basement membrane (Sherwood and Sternberg, 2003). The anchor cell can pass by the vulE cells and locate the vulF cells. The signal from the vulval to the invading AC acts at a distance because a distant 1° VPC, indicated by expression of egl-17::yfp during the L3 stage, can attract extension of the anchor cell.
During the L4 larval stage, the vulval cells move towards the anchor cell, invaginate, fuse into seven toroids (see Cell fusion), and during the L4-to-adult molt evert. The epidermal cells of the vulva organize the egg-laying system.
The ability of the vulval invagination to form is regulated by eight sqv genes (sqv-1, sqv-2, sqv-3, sqv-4, sqv-5, sqv-6, sqv-7, and sqv-8; Herman et al., 1999:). Hermaprodites with decreased sqv function have only a rudimentary invagination (SQuashed Vulva phenotype). The sqv genes encodes enzymes of a glycosaminoglycan biosynthesis (Herman and Horvitz, 1999; Berninsone et al., 2001; Hwang and Horvitz, 2002a; Hwang and Horvitz, 2002b; Hwang et al., 2003a; Hwang et al., 2003b). The sqv genes are expressed broadly in the vulva with the exception of sqv-4, which is expressed in vulC, D, E and F (Hwang and Horvitz, 2002b).
Morphogenesis also involves cytoskeletal reorganization, and proteins such as the ARL2 ortholog EVL-20 (Antoshechkin and Han, 2002). The Rac proteins encoded by mig-2 and ced-10 (Kishore and Sundaram, 2002) are necessary for appropriate morphogenesis of the vulva. The different vul cells extend processes toward the developing vulval lumen, contact each other and fuse to form toroidal adult vulval cells (Sharma-Kishore et al. 1999). A plexin homolog (a semaphorin receptor) is expressed sequentially from vulF to vulA as the toroids fuse (Dalpe et al., 2005).
The vulval epithelium organizes the development of the muscles and neurons that mediate egg-laying (Li and Chalfie, 1990). This is most obviously seen in dig-1 mutants in which the gonad is shifted anteriorly and the vulva, vulval muscles and neurons are all similarly shifted anteriorly (Thomas et al., 1990). Specific gene products expressed in subsets of vulval cells direct details of development of individual neurons. bam-2 is necessary for axonal branch termination of the VC motoneurons, and is expressed in the vulF cells (Colavita and Tessier-Lavigne, 2003). syg-1 and syg-2 are required for synapse formation by the HSN neurons (Shen and Bargmann, 2003; Shen et al., 2004). syg-2 is expressed in vulE cells and syg-1 in the HSNs.
There is now a reasonable conceptual model for understanding how the pattern of VPCs is established, but many components remain to be identified and the precise relationship among many of the genes has not been established. By contrast, there is only a rudimentary model for how the pattern of the seven vulval cell types are established, and even less of a picture of how the vulval cells undergo morphogenesis to form the adult structure. Similarly, our understanding of the control of timing of vulval development is also incomplete, and the relationship of the cell cycle to fate specification remains intriguing. In spite of these limitations, the extensive knowledge of C. elegans vulval development makes it an interesting case for computational modeling studies (e.g., Fisher et al., 2005). The precision of the wild-type pattern of VPC fates stands in stark contrast to the variable phenotypes of many mutants and perturbations; I view these phenotypes as providing a rich source of information about the underlying mechanisms and their dynamics.
I am grateful to all my colleagues over the past 25 years who have studied vulval development or at least been willing to discuss it, especially Bob Horvitz, John Sulston, Judith Kimble, Chip Ferguson, Iva Greenwald, Victor Ambros, John White, Bill Fixsen, Jim Thomas, Tim Schedl, Stuart Kim, Meera Sundaram, Hitoshi Sawa, Alex Hajnal, Ira Herskowitz, Geraldine Seydoux, David Eisenmann, and in my laboratory, Raffi Aroian, Russell Hill, Gregg Jongeward, Min Han, Hiroyuki Mori, Andy Golden, Paul Kayne, Linda Huang, Helen Chamberlin, Jane Mendel, Junho Lee, Kyria Tietze, Robert Palmer, Wendy Katz, Ralf Sommer, Anna Newman, Giovanni Lesa, Jing Liu, Tom Clandinin, Marie-Anne Felix, Keith Brown, Minqin Wang, Pheobe Tzou, Yvonne Hajdu-Cronin, Chris Lacenere, Charles Yoon, Neil Hopper, Chieh Chang, Nadeem Moghal, Rene Garcia, Aidyl Gonzalez-Serricchio, Martha Kirouac, Shahla Gharib, Bhagwati Gupta, David Sherwood, Byung Hwang, Takao Inoue, Cheryl Van Buskirk, Jolene Fernandes, Ted Ririe, David Goulet, and Edo Israeli. I thank Cheryl Van Buskirk and Iva Greenwald for helpful editorial comments.
I am grateful to the following sources that funded vulval development research in my laboratory: Searle Foundation (1988-1991); National Institutes of Health HD23690 (1988 -2000), March of Dimes (1988-1994), National Science Foundation (1988-1991), U.S. Army Breast Cancer Research Program (1992-1997) and the Howard Hughes Medical Institute (1989-present), as well as postdoctoral fellowships from American Cancer Society, Human Frontier Science Program, Life Sciences Research Foundation, Lymphoma and Leukemia Society, Damon Runyon Cancer Research Foundation, California Breast Cancer Research Program, European Molecular Biology Organization, National Institutes of Health, U.S.A. I am an Investigator with the Howard Hughes Medical Institute.
Ambros, V. (1999). Cell cycle-dependent sequencing of cell fate decisions in Caenorhabditis elegans vulva precursor cells. Development 126, 1947–1956. Abstract
Antoshechkin, I., and Han M. (2002). The C. elegans evl-20 gene is a homolog of the small GTPase ARL2 and regulates cytoskeleton dynamics during cytokinesis and morphogenesis. Dev. Cell 2, 579–591. Abstract Article
Aroian R.V., and Sternberg, P.W. (1991). Multiple functions of let-23, a Caenorhabditis elegans receptor tyrosine kinase gene required for vulval induction. Genetics 128, 251–267. Abstract
Battu, G., Hoier E.F., and Hajnal, A. (2003). The C. elegans G-protein-coupled receptor SRA-13 inhibits RAS/MAPK signalling during olfaction and vulval development. Development 130, 2567–2577. Abstract Article
Beitel, G.J., Clark, S.G., and Horvitz, H.R. (1990). Caenorhabditis elegans ras gene let-60 acts as a switch in the pathway of vulval induction. Nature 348, 503–509. Abstract Article
Beitel, G.J., Tuck, S., Greenwald, I., and Horvitz, H.R. (1995). The Caenorhabditis elegans gene lin-1 encodes an ETS-domain protein and defines a branch of the vulval induction pathway. Genes Dev. 9, 3149–3162. Abstract Article
Berninsone, P., Hwang, H.Y., Zemtseva, I., Horvitz, H.R., and Hirschberg, C.B. (2001). SQV-7, a protein involved in Caenorhabditis elegans epithelial invagination and early embryogenesis, transports UDP-glucuronic acid, UDP-N- acetylgalactosamine, and UDP-galactose. Proc. Natl. Acad. Sci. USA 98, 3738–3743. Abstract Article
Berset T., Hoier, E.F., Battu, G., Canevascini, S., and Hajnal, A. (2001). Notch inhibition of RAS signaling through MAP kinase phosphatase LIP-1 during C. elegans vulval development. Science 291, 1055–1058. Abstract Article
Boxem, M., Srinivasan, D.G., and van den Heuvel, S. (1999). The Caenorhabditis elegans gene ncc-1 encodes a cdc2-related kinase required for M phase in meiotic and mitotic cell divisions, but not for S phase. Development 126, 2227–2239. Abstract
Boxem, M., and van den Heuvel, S. (2001). lin-35 Rb and cki-1 Cip/Kip cooperate in developmental regulation of G1 progression in C. elegans. Development 128, 4349–4359. Abstract
Boxem, M., and van den Heuvel, S. (2002). C. elegans class B synthetic multivulva genes act in G(1) regulation. Curr. Biol. 12, 906–911. Abstract Article
Broday, L., Kolotuev, I., Didier, C., Bhoumik, A., Gupta, B.P., Sternberg, P.W., Podbilewicz, B., and Ronai, Z. (2004). The small ubiquitin-like modifier (SUMO) is required for gonadal and uterine-vulval morphogenesis in Caenorhabditis elegans. Genes Dev. 18, 2380–2391. Abstract Article
Bruinsma, J.J., Jirakulaporn, T., Muslin, A.J., and Kornfeld, K. (2002). Zinc ions and cation diffusion facilitator proteins regulate Ras-mediated signaling. Dev. Cell 2, 567–578. Abstract Article
Bulik, D.A., and Robbins, P.W. (2002). The Caenorhabditis elegans sqv genes and functions of proteoglycans in development. Biochim. Biophys. Acta 1573, 247–57.
Bulik, D.A., Wei, G., Toyoda, H., Kinoshita-Toyoda, A., Waldrip, W.R., Esko, J.D., Robbins, P.W., and Selleck, S.B. (2000). sqv-3, -7, and -8, a set of genes affecting morphogenesis in Caenorhabditis elegans, encode enzymes required for glycosaminoglycan biosynthesis. Proc. Natl. Acad. Sci. USA 97, 10838–10843. Article
Burdine, R.D., Branda, C.S., and Stern, M.J. (1998). EGL-17(FGF) expression coordinates the attraction of the migrating sex myoblasts with vulval induction in C. elegans. Development 125, 1083–1093. Abstract
Ceol, C.J., Horvitz, H.R. dpl-1 DP and efl-1 E2F act with lin-35 Rb to antagonize Ras signaling in C. elegans vulval development. Mol. Cell 7, 461–473. Article
Chamberlin, H.M., Palmer, R.E., Newman, A.P, Sternberg, P.W., Baillie DL, and Thomas, J.H. (1997). The PAX gene egl-38 mediates developmental patterning in Caenorhabditis elegans. Development 124, 3919–3928. Abstract
Chang, C., Hopper, N.A., and Sternberg, P.W. (2000). Caenorhabditis elegans SOS-1 is necessary for multiple RAS-mediated developmental signals. EMBO J. 19, 3283–3294. Abstract Article
Chang, C., Newman, A.P., and Sternberg, P.W. (1999). Reciprocal EGF signaling back to the uterus from the induced C. elegans vulva coordinates morphogenesis of epithelia. Curr. Biol. 9, 237–246. Abstract Article
Chen, N., and Greenwald, I. (2004). The lateral signal for LIN-12/Notch in C. elegans vulval development comprises redundant secreted and transmembrane DSL proteins. Dev. Cell 6, 183–192. Abstract Article
Cinar, H.N., Sweet, K.L, Hosemann, K.E., Earley, K., and Newman, A.P. (2001). The SEL-12 presenilin mediates induction of the Caenorhabditis elegans uterine pi cell fate. Dev. Biol. 237, 173–182. Abstract Article
Clandinin, T.R., Katz, W.S., and Sternberg, P.W. (1997). Caenorhabditis elegans HOM-C genes regulate the response of vulval precursor cells to inductive signal. Dev. Biol 182, 150–161. Abstract Article
Clark, S.H. (1992). Intercellular signalling and homeotic genes required during vulval development in C. elegans. Ph. D. Thesis, Massacusetts Institute of Technology.
Clark, S.G., Chisholm, A.D, and Horvitz, H.R. (1993). Control of cell fates in the central body region of C. elegans by the homeobox gene lin-39. Cell 74, 43–55. Abstract Article
Clark, S.G., Lu, X., and Horvitz, H.R. (1994). The Caenorhabditis eleganslocus lin-15, a negative regulator of a tyrosine kinase signaling pathway, encodes two different proteins. Genetics 137, 987–997. Abstract
Clark, S.G., Stern, M.J., and Horvitz, H.R. (1992). C. elegans cell-signalling gene sem-5 encodes a protein with SH2 and SH3 domains. Nature 356, 340–344. Abstract Article
Clark, S.G., Stern, M.J., and Horvitz, H.R. (1992). Genes involved in two Caenorhabditis elegans cell-signaling pathways. Cold Spring Harb. Symp. Quant. Biol. 57, 363–373. Abstract
Colavita, A., and Tessier-Lavigne, M. (2003). A Neurexin-related protein, BAM-2, terminates axonal branches in C. elegans. Science 302, 293–296. Abstract Article
Cui, M., Fay, D.S, and Han, M. (2004). lin-35/Rb cooperates with the SWI/SNF complex to control Caenorhabditis elegans larval development. Genetics 167, 1177–1185. Abstract Article
Cui, M., and Han, M. (2003). Cis regulatory requirements for vulval cell-specific expression of the Caenorhabditis elegans fibroblast growth factor gene egl-17. Dev. Biol. 257, 104–116. Abstract Article
Dalpe, G., Brown, L., and Culotti, J.G. (2005). Vulva morphogenesis involves attraction of plexin 1-expressing primordial vulva cells to semaphorin 1a sequentially expressed at the vulva midline. Development 132, 1387–1400. Article
Deshpande, R., Inoue, T., Priess, J.R., and Hill, R.J. (2005). lin-17/Frizzled and lin-18 regulate POP-1/TCF-1 localization and cell type specification during C. elegans vulval development. Dev. Biol. 278, 118–129. Article
Dutt, A., Canevascini, S., Froehli-Hoier, E., and Hajnal, A. (2004). EGF signal propagation during C. elegans vulval development mediated by ROM-1 rhomboid. PLoS Biol. 2, E334. Abstract Article
Eisenmann, D.M., and Kim, S.K. (2000). Protruding vulva mutants identify novel loci and Wnt signaling factors that function during Caenorhabditis elegans vulva development. Genetics 156, 1097–1116. Abstract
Eisenmann, D.M., Maloof, J.N., Simske, J.S., Kenyon, C., and Kim, S.K. (1998). The beta-catenin homolog BAR-1 and LET-60 Ras coordinately regulate the Hox gene lin-39 during Caenorhabditis elegans vulval development. Development 125, 3667–3680. Abstract
Euling, S., and Ambros, V. (1996). Reversal of cell fate determination in Caenorhabditis elegans vulval development. Development 122, 2507–2515. Abstract
Euling, S., and Ambros, V. (1996). Heterochronic genes control cell cycle progress and developmental competence of C. elegans vulva precursor cells. Cell 8, 667–676. Article
Fay, D.S., and Han, M. (2000). Mutations in cye-1, a Caenorhabditis elegans cyclin E homolog, reveal coordination between cell-cycle control and vulval development. Development 127, 4049–4060. Abstract
Ferguson, E.L., and Horvitz, H.R. (1985). Identification and characterization of 22 genes that affect the vulval cell lineages of the nematode Caenorhabditis elegans. Genetics 110, 17–72. Abstract
Ferguson, E.L., and Horvitz, H.R. (1989). The multivulva phenotype of certain Caenorhabditis elegans mutants results from defects in two functionally redundant pathways. Genetics 123, 109–121. Abstract
Ferguson, E.L., Sternberg, P.W., and Horvitz, H.R. (1987). A genetic pathway for the specification of the vulval cell lineages of Caenorhabditis elegans. Nature 326, 259–67. Erratum in: Nature 327, 82. Abstract Article
Fisher, J., Piterman, N., Hubbard, E.J., Stern, M.J., Harel, D. (2005). Computational insights into Caenorhabditis elegans vulval development. Proc. Natl. Acad. Sci. USA 102, 1951–1956. Article
Freyd, G., Kim, S.K., and Horvitz, H.R. (1990). Novel cysteine-rich motif and homeodomain in the product of the Caenorhabditis elegans cell lineage gene lin-11. Nature 344, 76–879. Abstract
Gleason, J.E., Korswagen, H.C., and Eisenmann, D.M. (2002). Activation of Wnt signaling bypasses the requirement for RTK/Ras signaling during C. elegans vulval induction. Genes Dev. 16, 1281–1290. Abstract Article
Grant, K.A., Hanna-Rose, W., and Han, M. (2000). sem-4 promotes vulval cell fate determination in Caenorhabditis elegans through regulation of lin-39 hox. Dev. Biol. 224, 496–506. Abstract Article
Greenwald, I., and Seydoux, G. (1990). Analysis of gain-of-function mutations of the lin-12 gene of Caenorhabditis elegans. Nature 346, 197–199. Article
Greenwald, I.S., Sternberg, P.W., and Horvitz, H.R. (1983). The lin-12 locus specifies cell fates in Caenorhabditis elegans. Cell 34, 435–444. Abstract Article
Gupta, B.P., and Sternberg, P.W. (2002). Tissue-specific regulation of the LIM homeobox gene lin-11 during development of the Caenorhabditis elegans egg-laying system. Dev. Biol. 247, 102–115. Abstract Article
Gupta, B.P., Wang, M., and Sternberg, P.W. (2003). The C. elegans LIM homeobox gene lin-11 specifies multiple cell fates during vulval development. Development 130, 2589–2601. Abstract Article
Hajnal, A. (2002). Fine-tuning the RAS signaling pathway: Zn(2+) makes the difference. Mol. Cell 9, 927–928. Article
Hajnal, A., Whitfield, C.W., Kim, S.K.. (1997). Inhibition of Caenorhabditis elegans vulval induction by gap-1 and by let-23 receptor tyrosine kinase. Genes Dev. 11, 2715–2728. Abstract
Han, M., Aroian, R.V., and Sternberg, P.W. (1990). The let-60 locus controls the switch between vulval and nonvulval cell fates in Caenorhabditis elegans. Genetics 126, 899–913.
Han, M., Golden, A., Han, Y., and Sternberg, P.W. (1993). C. elegans lin-45 raf gene participates in let-60 ras-stimulated vulval differentiation. Nature 363, 133–140. Abstract Article
Han, M., and Sternberg, P.W. (1990). let-60, a gene that specifies cell fates during C. elegans vulval induction, encodes a ras protein. Cell 63, 921–931. Abstract Article
Han, M., and Sternberg, P.W. (1991). Analysis of dominant-negative mutations of the Caenorhabditis elegans let-60 ras gene. Genes Dev. 5, 2188–2198.
Hanna-Rose, W., and Han, M. (1999). COG-2, a sox domain protein necessary for establishing a functional vulval-uterine connection in Caenorhabditis elegans. Development 126, 169–179. Abstract
Hanna-Rose, W., and Han, M. (2002). The Caenorhabditis elegans EGL-26 protein mediates vulval cell morphogenesis. Dev. Biol. 241, 247–258. Abstract Article
Hedgecock, E.M., and Herman, R.K. (1995). The ncl-1 gene and genetic mosaics of Caenorhabditis elegans. Genetics 141, 989–1006. Abstract
Herman, R.K., and Hedgecock, E.M. (1990). Limitation of the size of the vulval primordium of Caenorhabditis elegans by lin-15 expression in surrounding hypodermis. Nature 348, 169–171. Abstract Article
Herman, T., Hartwieg, E., and Horvitz, H.R. (1999). sqv mutants of Caenorhabditis elegans are defective in vulval epithelial invagination. Proc. Natl. Acad. Sci. USA. 96, 968–973. Abstract Article
Herman, T., and Horvitz, H.R. (1999). Three proteins involved in Caenorhabditis elegansvulval invagination are similar to components of a glycosylation pathway. Proc. Natl. Acad. Sci. USA. 96, 974–979. Article
Hill, R.J., and Sternberg, P.W. (1992). The gene lin-3 encodes an inductive signal for vulval development in C. elegans. Nature 358, 470–476. Abstract Article
Hong, Y., Roy, R., and Ambros, V. (1998). Developmental regulation of a cyclin-dependent kinase inhibitor controls postembryonic cell cycle progression in Caenorhabditis elegans. Development 125, 3585–3597. Abstract
Hopper, N.A., Lee, J., and Sternberg, P.W. (2000). ARK-1 inhibits EGFR signaling in C. elegans. Mol. Cell 6, 65–75. Abstract Article
Horvitz, H.R., and Sternberg, P.W. (1991). Multiple intercellular signalling systems control the development of the Caenorhabditis elegans vulva. Nature 351, 535–541. Abstract Article
Horvitz, H.R., Sulston, J.E. (1980). Isolation and genetic characterization of cell-lineage mutants of the nematode Caenorhabditis elegans. Genetics 96, 435–454. Abstract
Hoskins, R., Hajnal, A.F., Harp, S.A., Kim, S.K. (1996). The C. elegans vulval induction gene lin-2 encodes a member of the MAGUK family of cell junction proteins. Development 122, 97–111. Abstract
Huang, L.S., Tzou, P., and Sternberg, P.W. (1994). The lin-15 locus encodes two negative regulators of Caenorhabditis elegans vulval development. Mol. Biol. Cell 5, 395–411. Abstract
Hwang, H.Y., and Horvitz, H.R. (2002a). The SQV-1 UDP-glucuronic acid decarboxylase and the SQV-7 nucleotide-sugar transporter may act in the Golgi apparatus to affect Caenorhabditis elegans vulval morphogenesis and embryonic development. Proc. Natl. Acad. Sci. USA 99, 14218–14223. Abstract Article
Hwang, H.Y., and Horvitz, H.R. (2002b). The Caenorhabditis elegans vulval morphogenesis gene sqv-4 encodes a UDP-glucose dehydrogenase that is temporally and spatially regulated. Proc. Natl. Acad. Sci. USA 99, 14224–14229. Abstract Article
Hwang, H.Y., Olson, S.K., Esko, J.D., and Horvitz, H.R. (2003a). Caenorhabditis elegans early embryogenesis and vulval morphogenesis require chondroitin biosynthesis. Nature 423, 439–443. Abstract Article
Hwang, H.Y., Olson, S.K., Brown, J.R., Esko, J.D., and Horvitz, H.R. (2003b). The Caenorhabditis elegans genes sqv-2 and sqv-6, which are required for vulval morphogenesis, encode glycosaminoglycan galactosyltransferase II and xylosyltransferase. J. Biol. Chem. 278, 11735–11738. Abstract Article
Hwang, B.J., and Sternberg, P.W. (2004). A cell-specific enhancer that specifies lin-3 expression in the C. elegans anchor cell for vulval development. Development 131, 143–151. Article
Inoue, T., Oz, H.S., Wiland, D., Gharib, S., Deshpande, R., Hill, R.J., Katz, W.S., and Sternberg, P.W. (2004). C. elegans LIN-18 is a Ryk ortholog and functions in parallel to LIN-17/Frizzled in Wnt signaling. Cell 118, 795–806. Abstract Article
Inoue, T., Sherwood, D.R., Aspock, G., Butler, J.A., Gupta, B.P., Kirouac, M., Wang, M., Lee, P.Y., Kramer, J.M., Hope, I., Burglin, T.R., and Sternberg, P.W. (2002). Gene expression markers for Caenorhabditis elegans vulval cells. Mech. Dev. 119, S203–S209. Abstract Article
Inoue, T., Wang, M., Ririe, T.O., Fernandes, J.S., and Sternberg, P.W. (2005). Transcriptional network underlying Caenorhabditis elegans vulval development. Proc. Natl. Acad. Sci. USA
Jacobs, D., Beitel, G.J., Clark, S.G., Horvitz, H.R., Kornfeld, K. (1998). Gain-of-function mutations in the Caenorhabditis elegans lin-1 ETS gene identify a C-terminal regulatory domain phosphorylated by ERK MAP kinase. Genetics 149, 1809–1822. Abstract
Jongeward, G.D., Clandinin, T.R., and Sternberg, P.W. (1995). sli-1, a negative regulator of let-23-mediated signaling in C. elegans. Genetics 139, 1553–1566. Abstract
Kaech, S.M., Whitfield, C.W., Kim, S.K. (1998). The LIN-2/LIN-7/LIN-10 complex mediates basolateral membrane localization of the C. elegans EGF receptor LET-23 in vulval epithelial cells. Cell 94, 761–771.
Katz, W.S., Hill, R.J., Clandinin, T.R., and Sternberg, P.W. (1995). Different levels of the C. elegans growth factor LIN-3 promote distinct vulval precursor fates. Cell 82, 297–307. Abstract Article
Katz, W.S., Lesa, G.M., Yannoukakos, D., Clandinin, T.R., Schlessinger, J., and Sternberg, P.W. (1996). A point mutation in the extracellular domain activates LET-23, the Caenorhabditis elegans epidermal growth factor receptor homolog. Mol. Cell Biol. 16, 529–537. Abstract
Kimble, J. (1981). Alterations in cell lineage following laser ablation of cells in the somatic gonad of Caenorhabditis elegans. Dev. Biol. 87, 286–300. Abstract Article
Kirouac, M., and Sternberg, P.W. (2003). cis-Regulatory control of three cell fate-specific genes in vulval organogenesis of Caenorhabditis elegans and C. briggsae. Dev. Biol. 257, 85–103. Abstract Article
Kishore, R.S., and Sundaram, M.V. (2002). ced-10 Rac and mig-2 function redundantly and act with unc-73 trio to control the orientation of vulval cell divisions and migrations in Caenorhabditis elegans. Dev. Biol. 241, 339–348. Article
Koga, M., and Ohshima, Y. (1995). Mosaic analysis of the let-23 gene function in vulval induction of Caenorhabditis elegans. Development 121, 2655–2666. Abstract
Koh, K., Bernstein, Y., and Sundaram, M.V. (2004). The nT1 translocation separates vulval regulatory elements from the egl-18 and elt-6 GATA factor genes. Dev. Biol. 267, 252–263. Abstract Article
Koh, K., Peyrot, S.M., Wood, C.G., Wagmaister, J.A., Maduro, M.F., Eisenmann, D.M., and Rothman, J.H. (2002). Cell fates and fusion in the C. elegans vulval primordium are regulated by the EGL-18 and ELT-6 GATA factors, apparent direct targets of the LIN-39 Hox protein. Development 129, 5171–5180. Abstract
Kwon, J.Y., and Lee, J. (2001). Biological significance of a universally conserved transcription mediator in metazoan developmental signaling pathways. Development 128, 3095–3104. Abstract
Lackner, M.R., Kornfeld, K., Miller, L.M., Horvitz, H.R., Kim, S.K. (1994). A MAP kinase homolog, mpk-1, is involved in ras-mediated induction of vulval cell fates in Caenorhabditis elegans. Genes Dev. 8, 160–173.
Lee, J., Jongeward, G.D., and Sternberg, P.W. (1994). unc-101, a gene required for many aspects of Caenorhabditis elegans development and behavior, encodes a clathrin-associated protein. Genes Dev. 8, 60–73. Abstract
Lesa, G.M., and Sternberg, P.W. (1997). Positive and negative tissue-specific signaling by a nematode epidermal growth factor receptor. Mol. Biol. Cell 8, 779–793.
Levitan, D., and Greenwald, I. (1998). LIN-12 protein expression and localization during vulval development in C. elegans. Development 125, 3101–3109.
Li, C., and Chalfie, M. (1990). Organogenesis in C. elegans: positioning of neurons and muscles in the egg-laying system. Neuron 4, 681–695. Article
Lu, X., Horvitz, H.R. (1998). lin-35 and lin-53, two genes that antagonize a C. elegans Ras pathway, encode proteins similar to Rb and its binding protein RbAp48. Cell 95, 981–991. Abstract Article
Maloof, J.N., Kenyon, C. (1998). The Hox gene lin-39 is required during C. elegans vulval induction to select the outcome of Ras signaling. Development 125, 181–190. Abstract
Melendez, A., and Greenwald, I. (2000). Caenorhabditis elegans lin-13, a member of the LIN-35 Rb class of genes involved in vulval development, encodes a protein with zinc fingers and an LXCXE motif. Genetics 155, 1127–1137. Abstract
Miller, L.M., Gallegos, M.E., Morisseau, B.A., Kim, S.K. (1993). lin-31, a Caenorhabditis elegans HNF-3/fork head transcription factor homolog, specifies three alternative cell fates in vulval development. Genes Dev. 7, 933–947.
Miller, L.M., Hess, H.A., Doroquez, D.B., and Andrews, N.M. (2000). Null mutations in the lin-31 gene indicate two functions during Caenorhabditis elegans vulval development. Genetics 156, 1595–1602. Abstract
Moghal, N., Garcia, L.R., Khan, L.A., Iwasaki, K., and Sternberg, P.W. (2003). Modulation of EGF receptor-mediated vulva development by the heterotrimeric G-protein Galphaq and excitable cells in C. elegans. Development 130, 4553–4566. Abstract Article
Mohler, W.A., Shemer, G., del Campo, J.J., Valansi, C., Opoku-Serebuoh, E., Scranton, V., Assaf, N., White, J.G., Podbilewicz, B. (2002). Type I membrane protein EFF-1 is essential for developmental cell fusion. Dev. Cell 2, 355–362. Abstract Article
Myers, T.R., Greenwald, I. (2005). lin-35 Rb acts in the major hypodermis to oppose ras-mediated vulval induction in C. elegans. Dev. Cell. 8, 117–123. Abstract Article
Newman, A.P., Acton, G.Z., Hartwieg, E., Horvitz, H.R., and Sternberg, P.W. (1999). The lin-11 LIM domain transcription factor is necessary for morphogenesis of C. elegans uterine cells. Development 126, 5319–5326. Abstract
Newman, A.P., and Sternberg, P.W. (1996). Coordinated morphogenesis of epithelia during development of the Caenorhabditis elegans uterine-vulval connection. Proc. Natl. Acad. Sci. USA 93, 9329–9333. Abstract Article
Newman, A.P., Inoue, T., Wang, M., and Sternberg, P.W. (1999). The Caenorhabditis elegans heterochronic gene lin-29 coordinates the vulval-uterine-epidermal connections. Curr. Biol. 10, 1479–1488. Abstract Article
Newman, A.P., White, J.G., and Sternberg, P.W. (1995). The Caenorhabditis elegans lin-12 gene mediates induction of ventral uterine specialization by the anchor cell. Development 121, 263–271. Abstract
Newman, A.P., White, J.G., and Sternberg, P.W. (1996). Morphogenesis of the C. elegans hermaphrodite uterus. Development 122, 3617–3626. Abstract
Palmer, R.E., Inoue, T., Sherwood, D.R., Jiang, L.I., and Sternberg, P.W. (2002). Caenorhabditis elegans cog-1 locus encodes GTX/Nkx6.1 homeodomain proteins and regulates multiple aspects of reproductive system development. Dev. Biol. 252, 202–213. Abstract Article
Romagnolo, B., Jiang, M., Kiraly, M., Breton, C., Begley, R., Wang, J., Lund, J., Kim, S.K. (2002). Downstream targets of let-60 Ras in Caenorhabditis elegans. Dev. Biol. 247, 127–136. Abstract Article
Salser, S.J., Loer, C.M., Kenyon, C. (1993). Multiple HOM-C gene interactions specify cell fates in the nematode central nervous system. Genes Dev. 7, 1714–1724. Abstract
Sawa, H., Lobel, L., and Horvitz, H.R. (1996). The Caenorhabditis elegans gene lin-17, which is required for certain asymmetric cell divisions, encodes a putative seven-transmembrane protein similar to the Drosophila frizzled protein. Genes Dev. 10, 2189–2197. Abstract
Seydoux, G., Savage, C., and Greenwald, I. (1993). Isolation and characterization of mutations causing abnormal eversion of the vulva in Caenorhabditis elegans. Dev. Biol. 157, 423–436. Abstract Article
Sharma-Kishore, R., White, J.G., Southgate, E., and Podbilewicz, B. (1999). Formation of the vulva in Caenorhabditis elegans: a paradigm for organogenesis. Development 126, 691–699. Abstract
Shaye, D.D., and Greenwald, I. (2002). Endocytosis-mediated downregulation of LIN-12/Notch upon Ras activation in Caenorhabditis elegans. Nature 420, 686–690. Article
Shemer, G., Kishore, R., and Podbilewicz, B. (2000). Ring formation drives invagination of the vulva in Caenorhabditis elegans: Ras, cell fusion, and cell migration determine structural fates. Dev. Biol. 221, 233–248. Article
Shemer, G., and Podbilewicz, B. (2002). lin-39/Hox triggers cell division and represses EFF-1/fusogen-dependent vulval cell fusion. Genes Dev. 16, 3136–3141. Abstract Article
Shen, K., Bargmann, C.I. (2003). The immunoglobulin superfamily protein SYG-1 determines the location of specific synapses in C. elegans. Cell 112, 619–630. Abstract Article
Shen, K., Fetter, R.D., Bargmann, C.I. (2004). Synaptic specificity is generated by the synaptic guidepost protein SYG-2 and its receptor, SYG-1. Cell 116, 869–881. Erratum in: Cell (2004). 117, 553. Abstract Article
Sherwood, D.R., and Sternberg, P.W. (2003). Anchor cell invasion into the vulval epithelium in C. elegans. Dev. Cell 5, 21–31. Abstract Article
Shibata, Y., Branicky, R., Landaverde, I.O., Hekimi, S. (2003). Redox regulation of germline and vulval development in Caenorhabditis elegans. Science 302, 1779–1782. Abstract Article
Shim, J., and Sternberg, P.W., Lee, J. (2000). Distinct and redundant functions of mu1 medium chains of the AP-1 clathrin-associated protein complex in the nematode Caenorhabditis elegans. Mol. Biol. Cell 11, 2743–2756. Abstract
Simske, J.S., Kaech, S.M., Harp, S.A., Kim, S.K. (1996). LET-23 receptor localization by the cell junction protein LIN-7 during C. elegans vulval induction. Cell 85, 195–204. Article
Simske, J.S., and Kim, S.K. (1995). Sequential signalling during Caenorhabditis elegans vulval induction. Nature 375, 142–146. Abstract Article
Singh, N., and Han, M. (1995). sur-2, a novel gene, functions late in the let-60 ras-mediated signaling pathway during Caenorhabditis elegans vulval induction. Genes Dev. 9, 2251–65. Abstract
Solari, F., and Ahringer, J. (2000). NURD-complex genes antagonise Ras-induced vulval development in Caenorhabditis elegans. Curr. Biol. 10, 223–226. Abstract Article
Sternberg, P.W. (1988). Lateral inhibition during vulval induction in Caenorhabditis elegans. Nature 335, 551–554. Abstract Article
Sternberg, P.W. (2004). Developmental biology. A pattern of precision. Science 303, 637–638. Abstract Article
Sternberg, P.W., and and Horvitz, H.R. (1982). Postembryonic nongonadal cell lineages of the nematode Panagrellus redivivus: description and comparison with those of Caenorhabditis elegans. Dev. Biol. 93, 181–205. Abstract Article
Sternberg, P.W., and Horvitz, H.R. (1986). Pattern formation during vulval development in C. elegans. Cell 44, 761–772. Abstract Article
Sternberg, P.W., and Horvitz, H.R. (1988). lin-17 mutations of Caenorhabditis elegans disrupt certain asymmetric cell divisions. Dev. Biol. 130, 67–73. Abstract Article
Sternberg, P.W., and Horvitz, H.R. (1989). The combined action of two intercellular signaling pathways specifies three cell fates during vulval induction in C. elegans. Cell 58, 679–693. Abstract Article
Sulston, J.E., and Horvitz, H.R. (1977). Post-embryonic cell lineages of the nematode, Caenorhabditis elegans. Dev. Biol. 56, 110–156. Abstract Article
Sulston, J.E., and Horvitz, H.R. (1981). Abnormal cell lineages in mutants of the nematode Caenorhabditis elegans. Dev. Biol. 82, 41–55. Abstract Article
Sulston, J.E., White, J.G. (1980). Regulation and cell autonomy during postembryonic development of Caenorhabditis elegans. Dev. Biol. 78, 577–597. Abstract Article
Sundaram, M.V. (2004). Vulval development: the battle between Ras and Notch. Curr. Biol. 14, R311–R313. Abstract Article
Tan, P.B., Lackner, M.R., Kim, S.K. (1998). MAP kinase signaling specificity mediated by the LIN-1 Ets/LIN-31 WH transcription factor complex during C. elegans vulval induction. Cell 93, 569–580. Abstract Article
Tax, F.E., Thomas, J.H., Ferguson, E.L., and Horvitz, H.R. (1997). Identification and characterization of genes that interact with lin-12 in Caenorhabditis elegans. Genetics 147, 1675–1695. Abstract
Thomas, J.H., and Horvitz, H.R. (1999). The C. elegans gene lin-36 acts cell autonomously in the lin-35 Rb pathway. Development 126, 3449–3459. Abstract
Thomas, J.H., Stern, M.J., and Horvitz, H.R. (1990). Cell interactions coordinate the development of the C. elegans egg-laying system. Cell 62, 1041–1052. Abstract Article
Tuck, S., and Greenwald I. (1995). lin-25, a gene required for vulval induction in Caenorhabditis elegans. Genes Dev. 9, 341–357. Abstract
Wang, M., and Sternberg, P.W. (1999). Competence and commitment of Caenorhabditis elegans vulval precursor cells. Dev. Biol. 212, 12–24. Abstract Article
Wang, M., and Sternberg, P.W. (2000). Patterning of the C. elegans 1 degrees vulval lineage by RAS and Wnt pathways. Development 127, 5047–5058. Abstract
Yang, L., Sym, M., and Kenyon, C. (2005). The roles of two C. elegans HOX co-factor orthologs in cell migration and vulva development. Development 132, 1413–1428. Abstract Article
Yochem, J., Weston, K., and Greenwald, I. (1988). The Caenorhabditis elegans lin-12 gene encodes a transmembrane protein with overall similarity to Drosophila Notch. Nature 335, 547–550. Abstract Article
Yoda, A., Kouike, H., Okano, H., and Sawa, H. (2005). Components of the transcriptional Mediator complex are required for asymmetric cell division in C. elegans. Development 132, 1885–1893. Article
Yoder, J.H., Chong, H., Guan, K.L., Han, M. (2004). Modulation of KSR activity in Caenorhabditis elegans by Zn ions, PAR-1 kinase and PP2A phosphatase. EMBO J. 23, 111–119. Abstract Article
Yoo, A.S., Bais, C., and Greenwald, I. (2004). Crosstalk between the EGFR and LIN-12/Notch pathways in C. elegans vulval development. Science 303, 663–666. Abstract Article
Yoon, C.H., Chang, C., Hopper, N.A., Lesa, G.M., and Sternberg, P.W. (2000). Requirements of multiple domains of SLI-1, a Caenorhabditis elegans homologue of c-Cbl, and an inhibitory tyrosine in LET-23 in regulating vulval differentiation. Mol. Biol. Cell 11, 4019–4031. Abstract
Yoon, C.H., Lee, J., Jongeward, G.D., and Sternberg, P.W. (1995). Similarity of sli-1, a regulator of vulval development in C. elegans, to the mammalian proto-oncogene c-cbl. Science 269, 1102–1105. Abstract
*Edited by Barbara J. Meyer. Last revised June 18, 2005. Published June, 25 2005. This chapter should be cited as: Sternberg, P.W., Vulval development (June, 25 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.6.1, http://www.wormbook.org.
Copyright: © 2005 Paul W. Sternberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: pws@caltech.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.