Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Small GTPases of the Ras superfamily are key regulators of diverse cellular and developmental events, including differentiation, cell division, vesicle transport, nuclear assembly, and control of the cytoskeleton. The C. elegans genome encodes 56 members of the major Ras GTPase subfamilies, including the Ras/Ral/Rap family, the Rho family, the Rab family, Ran, and the Arf/Sar family. Studies in C. elegans have shown that Ras/Rap family members control cell fate specification and differentiation; Rho GTPases control morphogenesis and actin dynamics, including axon pathfinding and cell migration; Rab GTPases control synaptic vesicle trafficking and release and gene expression responses in innate immunity; the Ran GTPase controls nuclear import/export, nuclear reassembly after mitosis, and kinetechore association with microtubules; and Arf/Sar GTPases control morphogenesis and microtubule organization and possibly cilia development. Functions for many of the small GTPases remain to be discovered, and continuing studies in C. elegans will elucidate the roles of these molecules in animal development.
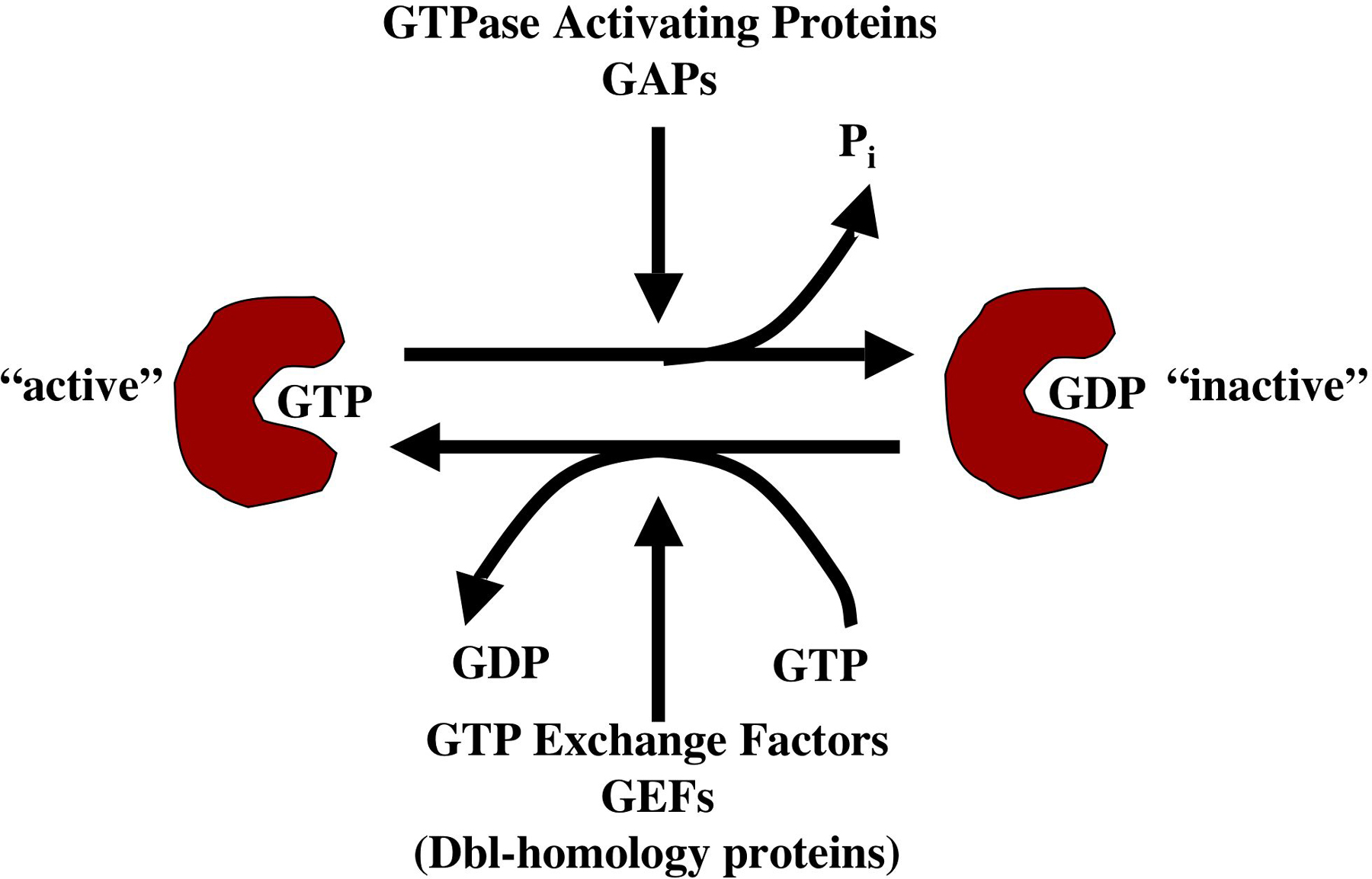
Small GTPases of the Ras superfamily are key regulators of diverse cellular and developmental events, including differentiation, cell division, vesicle transport, nuclear assembly, and control of the cytoskeleton. Many Ras-superfamily small GTPases are components of signaling pathways that link extracellular signals via transmembrane receptors to cytoplasmic or nuclear responses. Ras-superfamily small GTPases and their G-protein cousins act by a conserved mechanism (Figure 1): the molecules' signaling activities differ when bound to GTP versus GDP, and their GTPase activity (which hydrolyzes GTP to GDP), determines their GTP-bound versus GDP-bound state. Generally, GTP-bound molecules are thought to be active signaling molecules and GDP-bound forms are thought to be inactive (Bourne et al., 1991). Molecules that regulate Ras GTPases include GTPase activating proteins (GAPs), which promote the GDP-bound state of Ras GTPases by activating their GTPase activity and thus favor the inactive state, and GTP exchange factors (GEFs) which promote the active, GTP-bound state by facilitating the exchange of GDP for GTP.
|
Figure 1. The GTPase cycle. Ras-superfamily GTPases bound to GTP are active signaling molecules. The intrinsic GTPase activity of the molecule hydrolyzes GTP to GDP, and GDP-bound forms are inactive. GTPase activating proteins (GAPs) enhance GTPase activity and favor the inactive, GDP-bound state. GTP exchange factors (GEFs) faciltiate exchange of GDP with GTP and thus favor the active, GTP-bound state.
Canonical mutations that affect the GTPase cycle lead to constitutively-active or dominant-negative molecules (Bourne et al., 1991). Mutations that abolish GTPase activity (e.g., glycine 12 to valine; G12V) result in constitutive activation of the small GTPase, and mutations that presumably affect interaction of the GTPase with its effectors (e.g., threonine 17 to asparagine; T17N) result in dominant-negative molecules. Both activated and dominant- negative GTPases can dominantly perturb processes in which the GTPases are involved.
The C. elegans genome encodes 56 members of the major Ras GTPase subfamilies, including the Ras/Ral/Rap family, the Rho family, the Rab family, Ran, and the Arf/Sar family (Table 1). Furthermore, the C. elegans genome encodes GEFs and GAPs for the distinct GTPase families. For example, C. elegans has 19 genes that encode Rho GEFs of the Dbl-homology family, and 20 genes that encode Rho-family GAPs. Much is known about the roles of Ras and Rho family GTPases in C. elegans development, while many other GTPase families (e.g., Rab) remain uncharacterized. Key to understanding small GTPase function in development will be to determine the differential roles of GTPases and their GEFs, GAPs, and other signaling partners in different developmental events, and ongoing studies in C. elegans will contribute to this goal.
Table 1. Ras superfamily GTPases in C. elegans †
| C. elegans gene designation a | C. elegans gene name b | Human/mouse homolog c |
|---|---|---|
| Ras/Rap/Ral family | ||
| ZK792.6 | let-60 | K-Ras |
| F17C8.4 | ras-2 | M-Ras |
| C44C11.1 | ras-1 | R-Ras |
| C54A12.4 | – | Di-Ras2 |
| C27B7.8 | rap-1 | Rap-1b |
| C25D7.7 | rap-2 | Rap-2a |
| C08F8.7 | rap-3 | Rap-1b |
| T26C12.3 | – | Rap-2c |
| F54C8.5 | – | RheB |
| Y53G8AR.3 | – | Ral-A |
| C14A11.7 | ssr-1 | Dexamethasone-induced Ras-related protein 1 |
| Rho family | ||
| Y51H4A.3 | rho-1 | RhoA |
| R07G3.1 | cdc-42 | Cdc42 |
| C09G12.8 | ced-10 | Rac1 |
| C35C5.4 | mig-2 | RhoG d |
| F22E12.2 | Wrch1 | |
| Y32F6B.3 | conserved Cdc42-like P60953 | |
| Rab family | ||
| C39F7.4 | rab-1 | Rab1 |
| F53F10.4 | rab-2/unc-108 | Rab2 |
| C18A3.6 | rab-3 | Rab3 |
| F26H9.6 | rab-5 | Rab5 |
| F59B2.7 | rab-6.1 | Rab6 |
| T25G12.4 | rab-6.2 | Rab6 |
| D1037.4 | rab-8 | Rab8 |
| T23H2.5 | rab-10 | Rab10 |
| F53G12.1 | rab-11.1 | Rab11 |
| W04G5.2 | rab-11.2 | Rab11 |
| K09A9.2 | rab-14 | Rab14 |
| Y92C3B.3 | rab-18 | Rab18 |
| Y62E10A.9 | rab-19 | Rab19 |
| T01B7.3 | rab-21 | Rab21 |
| Y87G2A.4 | rab-27 | Rab27 |
| Y11D7A.4 | rab-28 | Rab28 |
| Y45F3A.2 | rab-30 | Rab30 |
| F43D9.2 | rab-33 | Rab33 |
| Y47D3A.25 | rab-35 | Rab35 |
| W01H2.3 | rab-37 | Rab37 |
| C56E6.2 | – | Rab6 |
| F11A5.3 | – | Rab2 |
| Arf/Sar family | ||
| ZK632.8 | arl-5 | Arf8 |
| B0336.2 | arf-1.2 | Arf1 |
| C38D4.8 | arl-6 | Arf6 |
| F54C9.10 | arl-1 | Arf1 |
| ZK180.4 | – | Sar1b |
| F45E4.1 | arf-1.1 | Arf1 |
| Y57G11C.13 | – | FLJ10702 Arf-family |
| F19H8.3 | arl-3 | Arf3 |
| F57H12.1 | arf-3 | Arf5 |
| Y116A8C.12 | arf-6 | Arf6 |
| Y37E3.5 | – | Arf2 |
| F22B5.1 | evl-20 | Arf2 |
| Ran family | ||
| K01G5.4 | ran-1 | Ran |
|
aThe C. elegans gene designation as noted on Wormbase. bThe three-letter C. elegans gene designation. Some genes have not yet been assigned a three letter designation. cSimilarities are as noted in Wormbase. dThe mouse and human genomes do not include genes similar to Mtl Rac (i.e. Racs with an N-terminal myristoylation sequence). However, vertebrate RhoG might be the functional equivalent of Mtl Rac (DeBakker et al., 2004). |
||
C. elegans has 11 members of the Ras/Rap/Ral family of small GTPases. The roles of many of these molecules in C. elegans development remain mysterious. The K-Ras homolog and the Rap subfamily have been analyzed.
The let-60 gene encodes the C. elegans K-Ras homolog (Beitel et al., 1990; Han and Sternberg, 1990). let-60 has been studied extensively in vulval cell fate induction (Beitel et al., 1990; Ferguson and Horvitz, 1985; Ferguson et al., 1987; Han et al., 1990). let-60 Ras affects other developmental events, including germ cell meiotic maturation (Church et al., 1995) and migration of the sex myoblast cells that produce the muscles that control vulval opening (Clark et al., 1992; Stern et al., 1993; Sundaram et al., 1996). A detailed description of let-60 function in C. elegans development can be found in RTKRas/MAP kinase signaling.
C. elegans harbors three Rap-like genes. rap-1 and rap-2 are very similar to vertebrate Rap-1b and Rap-2a. rap-3 shows less similarity to vertebrate Rap genes but is most similar to Rap-1b. In vertebrates and Drosophila, Rap proteins have been implicated in immune function, cell cycle control, Ras regulation, cell adhesion, and cytoskeletal rearrangements (Bos and Zwartkruis, 1999; Price et al., 2004; Rangarajan et al., 2003). Rap function in C. elegans involves the function and morphogenesis of hypodermal cells but not their generation or specification (Berkel et al., 2005). rap-1 mutants display defects in hypodermal morphogenesis and function, including formation of adherens junctions, secretion of cuticle and basement membrane, and hypodermal cell integrity and viability. While rap-2 mutations on their own are wild-type, rap-2 mutations enhance the effects of rap-1, suggesting that rap-1 and rap-2 act redundantly and can partially compensate for each other's loss (Berkel et al., 2005).
Consistent with a role of rap-1 and rap-2 in hypodermal function, mutations in pxf-1, which encodes a Rap GEF similar to PDZ-GEF/RA GEF, cause a phenotype similar to rap-1; rap-2 double mutants (Berkel et al., 2005). Furthermore, constitutively-active RAP-1(G12V) can partially rescue the pxf-1 mutant phenotype, indicating that PXF-1 and RAP-1 act together to control hypodermal morphogenesis and function.
C. elegans has one Rho (rho-1), one Cdc42 (cdc-42), two canonical Racs similar to vertebrate Rac1 (ced-10 and rac-2), and one Mtl Rac (mig-2). Mtl Racs, defined by C. elegans MIG-2 and Drosophila Mtl, are similar to both Rac and Cdc42 but interact functionally with Racs. Mtl Racs contain an N-terminal myristoylation signal not found in other Racs, and vertebrates apparently do not have an equivalent GTPase. However, recent work indicates that RhoG might be the functional equivalent of Mtl Rac in vertebrates (deBakker et al., 2004). Other Rho-family GTPases encoded in the C. elegans genome include a Wrch1-like molecule (F22E12.2 in C. elegans), a Cdc42-like GTPase that controls cytoskeletal rearrangements downstream of Wnt signaling (Tao et al., 2001), and another conserved Cdc42-like GTPase of unknown function (Y32F6B.3 in C. elegans).
Rho GTPases are key regulators of actin cytoskeleton organization and cellular morphogenesis and have been analyzed extensively in C. elegans. While Rho GTPases each have unique roles in development, an emerging theme in Rho GTPase function is that Rho GTPases have redundant roles with other Rho GTPases in certain developmental processes (i.e. compensatory, overlapping roles), indicating that distinct Rho GTPases might control similar molecular and cellular processes.
To date no mutations of rho-1 have been identified. rho-1 function has been ascertained using RNAi and by transgenic expression of constitutively-active and dominant-negative forms of rho-1.
RNAi-mediated knockdown of rho-1 results in early embryonic arrest, often with a failure of cytokinesis (Jantsch-Plunger et al., 2000; Spencer et al., 2001). Embryos that arrested later in development displayed severe defects in tissue morphogenesis. Other mutations that affect embryonic morphogenesis and cytokinesis identified other genes that interact genetically with rho-1 in this process. let-502 encodes a Rho-binding kinase and mel-11 encodes a myosin phosphatase (Piekny and Mains, 2002; Wissmann et al., 1999; Wissmann et al., 1997). These molecules likely affect actin-myosin cytoskeletal dynamics of the contractile ring during cytokinesis in response to Rho signaling. The cyk-4 gene, identifed in a screen for mutations affecting embryonic cytokinesis, encodes a Rho GAP that stimulates GTPase activity of Rho, Rac, and Cdc42 and that likely modulates RHO-1 activity in cytokinesis (Jantsch-Plunger et al., 2000).
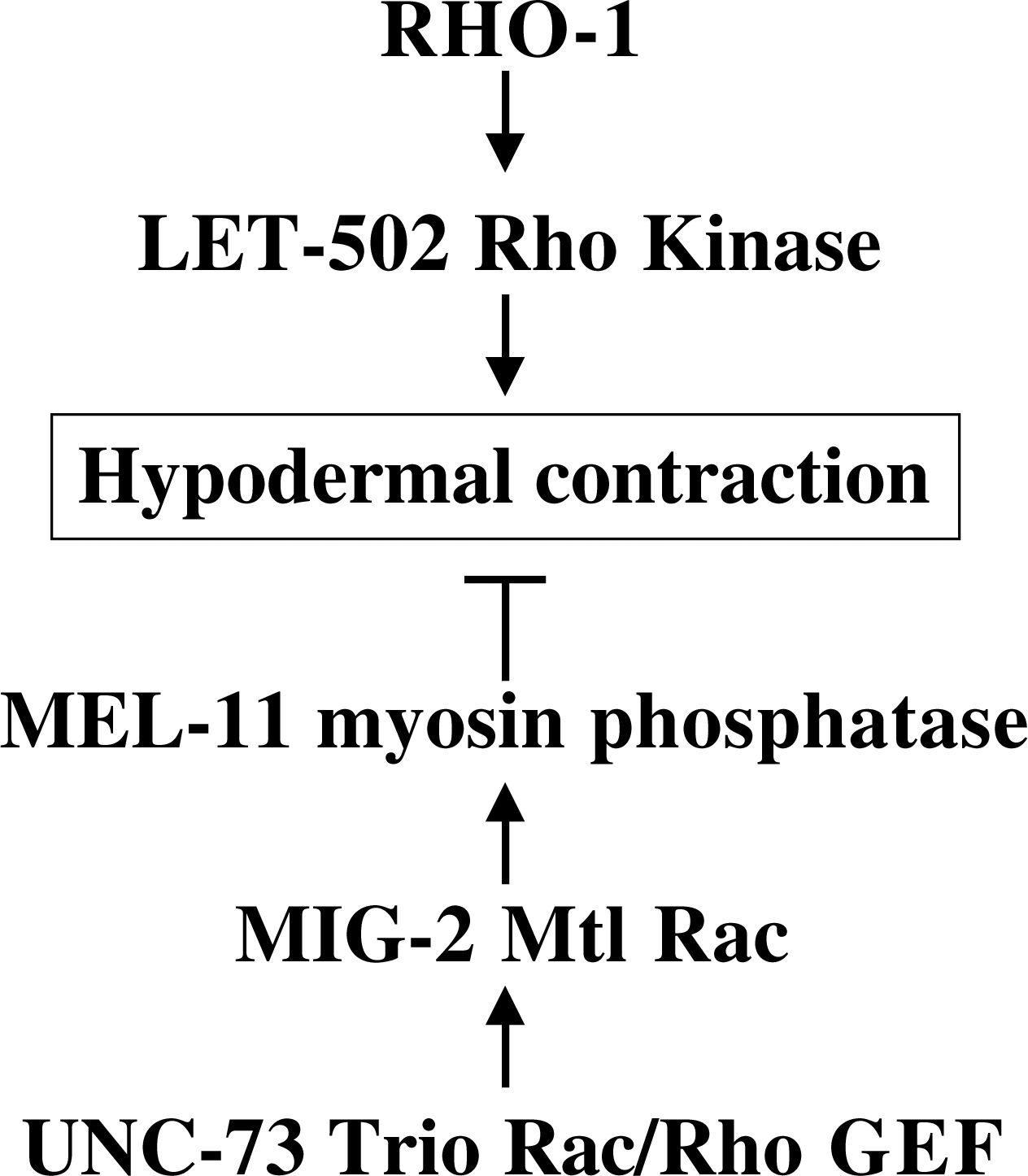
LET-502 and MEL-11 also control embryonic elongation. The hypodermal cells which epibolize the embryo contain circumferential rings of actin that contract, resulting in a "squeezing" of the embryo and a subsequent increase in length. LET-502 and MEL-11 have opposing roles in this process (LET-502 normally enhances contraction and MEL-11 normally inhibits contraction; Wissmann et al., 1999; Wissmann et al., 1997; Figure 2). Interestingly, the Mtl Rac GTPase MIG-2 and the UNC-73 Trio Rac/Rho GEF act in the MEL-11 pathway in this process. UNC-73 Trio acts as a GEF for RHO-1, CED-10, and MIG-2 (Spencer et al., 2001; Steven et al., 1998; Wu et al., 2002). These results indicate that MIG-2 Mtl Rac and RHO-1 might have opposing roles in embryonic elongation (Piekny et al., 2000; Figure 2).
|
Figure 2. The opposing roles of RHO-1 and MIG-2 Rac in hypdermal contraction during embryonic elongation. Circumferential actin bundles in the hypodermis contract to drive embryonic elongation. RHO-1 stimulates hypodermal contraction via the LET-502 Rho-binding kinase, and MIG-2 Mtl Rac inhibits hypodermal contraction via MEL-11 myosin phosphatase in response to UNC-73 Trio Rac/Rho GEF.
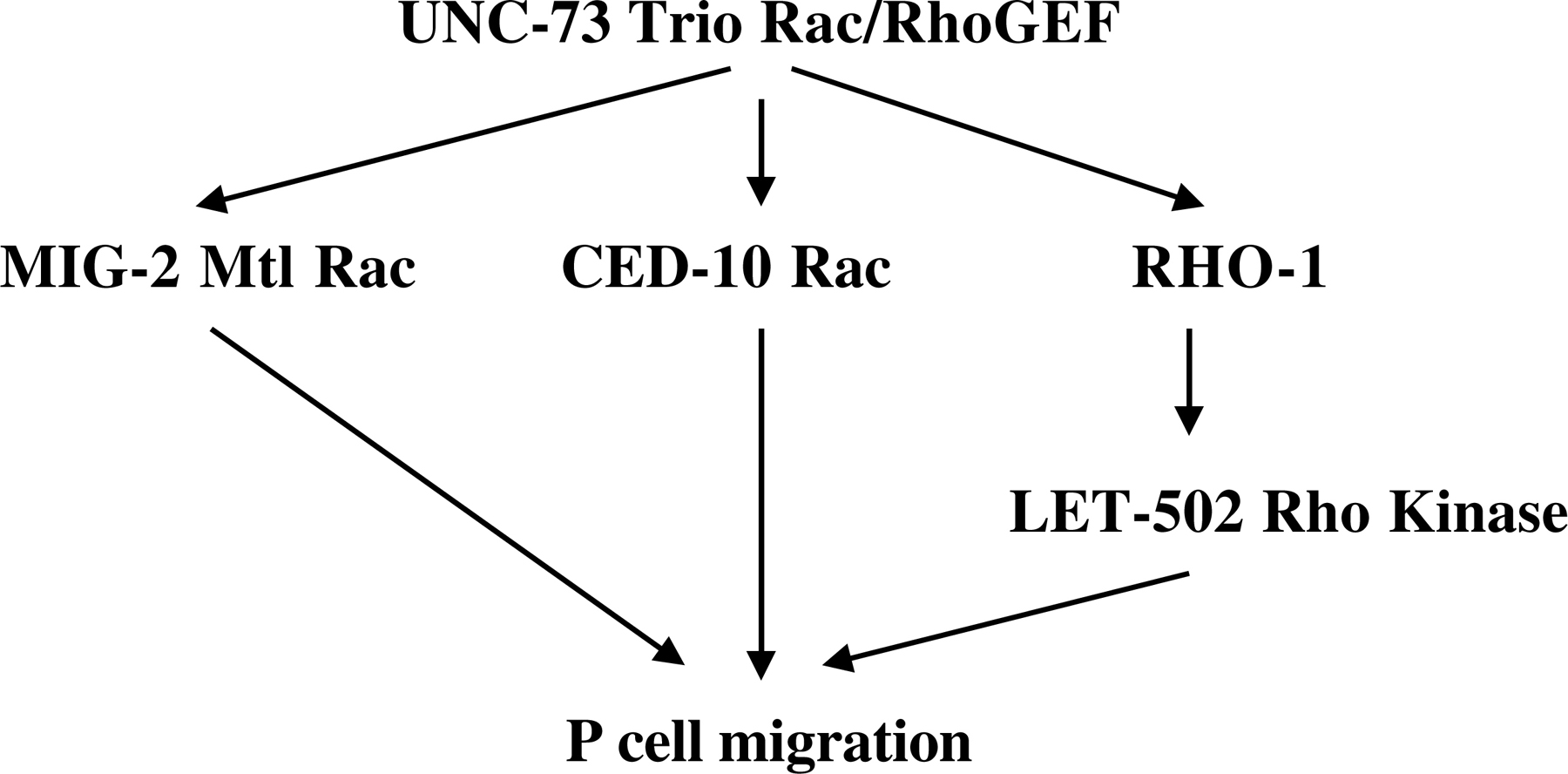
Later in development, rho-1 controls the ventral migrations of the P cells (Spencer et al., 2001). The P cell nuclei are born in sublateral locations and migrate ventrally to align at the ventral midline. RNAi of rho-1 and dominant-negative rho-1 transgenic expression perturb this migration of the P cells. The UNC-73 Trio Rac/Rho GEF acts upstream of RHO-1 in P cell migration (UNC-73 acts as a GEF on Rho and activated RHO-1 partially rescues an unc-73 mutant), and the LET-502 Rho-binding kinase acts downstream of RHO-1 in this process (a let-502 mutation is not rescued by activated rho-1; Spencer et al., 2001; Steven et al., 1998; Figure 3). Interestingly, the Rac GTPases CED-10 and MIG-2 control P-cell migration in a parallel redundant manner with RHO-1 (e.g., RHO-1 activity can partially compensate for loss of Rac activity in P cell migration; Spencer et al., 2001).
|
Figure 3. Overlapping roles of RHO-1, MIG-2 Mtl Rac and CED-10 Rac in P cell migration. Ventral P-cell migration is controlled by MIG-2 Mtl Rac, CED-10 Rac and RHO-1 in a redundant manner. UNC-73 Trio Rac/Rho GEF controls all three GTPases in P-cell migration, and LET-502 Rho-binding kinase acts downstream of RHO-1 in this event.
RHO-1 also controls neuronal cell shape after the establishment of the normal axon and dendrite morphology of the neuron. Expression of dominant-negative RHO-1(T19N) in the ASE sensory neuron resulted in an expanded cell body morphology and ectopic neurite initiation (Zallen et al., 2000). This phenotype is similar to that caused by loss of function of the SAX-1 Ndr kinase (Zallen et al., 2000), a known downstream target of Rho activity in other systems. RHO-1 and SAX-1 might act together to regulate neuronal morphogenesis.
RNAi of cdc-42 causes defects in embryonic cytokinesis similar to rho-1(RNAi) (Kay and Hunter, 2001). cdc-42(RNAi) also perturbs the polarity of the single-celled zygote and results in defects in anterior-posterior axis formation and mitotic spindle orientation (Gotta et al., 2001; Kay and Hunter, 2001). Upon fertilization, sperm entry results in a rearrangement of the zygote cytoplasm such that anterior cortex accumulates the PAR-3 and PAR-6 proteins and the posterior cortex accumulates the PAR-2 protein, which together define the anterior-posterior axis of the organism (Etemad-Moghadam et al., 1995; Hung and Kemphues, 1999; Levitan et al., 1994; Watts et al., 1996). The PAR proteins associate with the actin cytoskeleton at the cell cortex, and the actin cytoskeleton is required for their localization. As a result of PAR gene localization, the first cell division generates a larger anterior cell and a smaller posterior cell, whose spindle rotates 90° in relation to the anterior cell (Kemphues et al., 1988). Mutations in the par genes disrupt zygotic polarity and result in the symmetric division of zygote and defects in spindle orientation. RNAi of cdc-42 results in a similar loss of zygotic polarity and an associated failure to localize the PAR proteins to their proper domains (Gotta et al., 2001; Kay and Hunter, 2001). Thus, cdc-42 might control the localization of the PAR proteins or their interaction with the actin cytoskeleton. CDC-42 physically associates with PAR-6, a CRIB- and PDZ-domain-containing component of a conserved complex including PAR-3, and PKC-1, a protein kinase C ortholog, that defines the anterior cortical domain in the zygote (Gotta et al., 2001).
A screen for mutations that affect the development of the excretory cell identified exc-5, which encodes a putative Cdc42 GEF similar to the human faciogenital dysplasia-1 protein (Buechner et al., 1999; Gao et al., 2001; Suzuki et al., 2001). EXC-5 controls the formation of apical versus basolateral cell surfaces and cytoskeleton in the excretory canals: exc-5 loss of function mutants display large varicosities in the canals due to a failure of apical surface formation; and exc-5 overactivity results in foreshortened canals due to overproliferation of apical structures at the expense of basolateral structures. A direct role of CDC-42 in this process has not been demonstrated. However, the similarity of EXC-5 to the Cdc42 GEF FGD1 suggests that CDC-42 might be involved in excretory cell apical-basolateral polarity.
C. elegans WSP-1 is the homolog of the human Wiskcott-Aldrich syndrome protein (WASP). WASP is a downstream effector of Cdc42 that activates the Arp2/3 complex, an actin nucleating and branching complex. During C. elegans gastrulation, the dorsally-located hypodermal cells migrate toward the ventral midline where they meet and completely surround the embryo in a process known as ventral closure (Simske and Hardin, 2001; Sulston et al., 1983; Williams-Masson et al., 1997). C. elegans wsp-1 mutants display a low-penetrance embryonic lethality due to a failure of migration of the hypodermal cells and ventral closure (Withee et al., 2004).
Three Rac-like genes are present in the C. elegans genome. Two genes, ced-10 and rac-2, encode Rac1-like molecules, and mig-2 encodes an Mtl Rac (Lundquist et al., 2001; Reddien and Horvitz, 2000; Zipkin et al., 1997). ced-10 mutations were identified in mutant screens for defective apoptotic cell corpse phagocytosis (Ellis et al., 1991), and mig-2 mutations were identified in screens for cell migration-defective mutants (Zipkin et al., 1997). No mutations that perturb rac-2 function exist to date. rac-2 lof is induced using RNAi, and has no apparent phenotypic consequence except in combination with ced-10 and mig-2 mutations (see Overlapping roles of Racs in development).
Hypomorphic ced-10 mutations cause failure of phagocytosis of cell corpses after programmed cell death (Ellis et al., 1991; Reddien and Horvitz, 2000). Upon programmed death of a cell, neighboring cells extend actin-based plasma membrane protrusions to engulf and metabolize the cell corpse. ced-10 mutants fail in this process, resulting in the persistence of unengulfed cell corpses. ced-10 might be required to organize the actin cytoskeleton underlying the extension of cellular protrusions of the engulfing cell. Other engulfment mutations identified the ced-1, ced-2, ced-5, ced-6, ced-7, and ced-12 genes (Ellis et al., 1991). These genes can be placed into two pathways based upon redundancy of function in engulfment: mutations in ced-1, ced-6, and ced-7 enhance the engulfment phenotypes of ced-2, ced-5, and ced-10, whereas mutations within the two groups do not enhance each others' phenotypes. This suggests that two parallel pathways exist to control phagocytosis, one of which involves ced-10 Rac.
Molecular and genetic analyses revealed other CED-10 pathway members in phagocytosis (Figure 4), including CED-2, a CrkII-like receptor adapter protein (Reddien and Horvitz, 2000), and CED-5, a CDM-family Rac GEF (Wu and Horvitz, 1998). Furthermore, CED-12 is an ELMO-like protein, which cooperates with CDM proteins in Rac GTP exchange (Gumienny et al., 2001; Lu et al., 2004; Zhou et al., 2001).
|
Figure 4. Modularity of Rac signaling in different developmental processes. See text for details. (A) CED-10 Rac signaling in cell corpse phagocytosis. CED-10 Rac is controlled by CED-2 CrkII, CED-5 CDM and CED-12 ELMO in cell corpse phagocytosis. UNC-73 and MIG-2 act upstream of CED-12 and CED-5 in CED-10 activation. (B) CED-10 Rac is controlled by UNC-73 Trio Rac/Rho GEF in axon pathfinding. UNC-73 Trio has no apparent role in phagocytosis. CED-10 Rac might act downstream of the UNC-40 DCC receptor or the Integrin receptor. (C) In gastrulation, CED-10 Rac acts with GEX-2 Sra-1, GEX-3 Hem2, and possibly WVE-1 WAVE. The receptors that control Rac in phagocytosis and gastrulation, if any, are unknown.
Interestingly, MIG-2 Mtl Rac and the UNC-73 Trio Rac/Rho GEF act upstream of the CED-5/ CED-12/CED-10 complex in phagocytosis (deBakker et al., 2004; Figure 4). In response to UNC-73 GTP exchange, active GTP-bound MIG-2 interacts with CED-12 resulting in CED-5-mediated GTP exchange and activation of CED-10 (deBakker et al., 2004). Thus, Rac-like GTPases can act in a heirarchical fashion during phagocytosis.
ced-10 mutations also affect the migration of the gonadal distal tip cells, which results in misshapen gonad arms in ced-10 mutants (Lundquist et al., 2001). ced-5 and ced-12 also affect distal tip cell migration, as does the unc-73 Trio-like Rho/Rac GEF (which also acts with rho-1 in P-cell migration; Gumienny et al., 2001; Lundquist et al., 2001; Zhou et al., 2001).
Null alleles of ced-10 cause maternal-effect embryonic lethality with defects in early cell movements and gastrulation resulting in failure of the endoderm to be surrounded by the ectoderm (the gut on the exterior (Gex) phenotype; Lundquist et al., 2001; Soto et al., 2002). ced-10 might control actin rearrangements involved in movement and migration during gastrulation. Mutations in gex-1, gex-2 and gex-3 cause a similar maternal effect lethal phenotype. GEX-2 encodes an Sra-1-like molecule, and GEX-3 encodes a Hem2-like molecule. Sra-1 and Hem2, along with Nck, form a complex with the Arp2/3 complex activator WAVE (Eden et al., 2002). This complex might act downstream of CED-10 Rac in gastrulation (Figure 4).
Null mutations in mig-2 were isolated on the basis of defective migration of the Q cells and their daughters, and activated mig-2 alleles but not null mig-2 alleles affect CAN neuron migration (Zipkin et al., 1997). mig-2 mutations also display defective migration of the gonadal distal tip cells similar to ced-10 (Lundquist et al., 2001). The UNC-73 Trio Rac/Rho GEF acts with MIG-2 in both processes. As described for CED-10, MIG-2 and the UNC-73 Trio Rac/Rho GEF act upstream of the CED-12/CED-5/CED-10 complex to control phagocytosis (Figure 4).
Unlike rho-1 and cdc-42, which are the only members of their Rho subfamily encoded in the genome, three Rac genes, ced-10, mig-2 and rac-2, have both unique and redundant roles in diverse developmental events. While ced-10 and mig-2 mutations alone affect some developmental processes (e.g., phagocytosis and cell migration), analysis of pairwise Rac double mutant combinations revealed processes in which ced-10, mig-2 and rac-2 have overlapping and compensatory roles (i.e. in which they act redundantly).
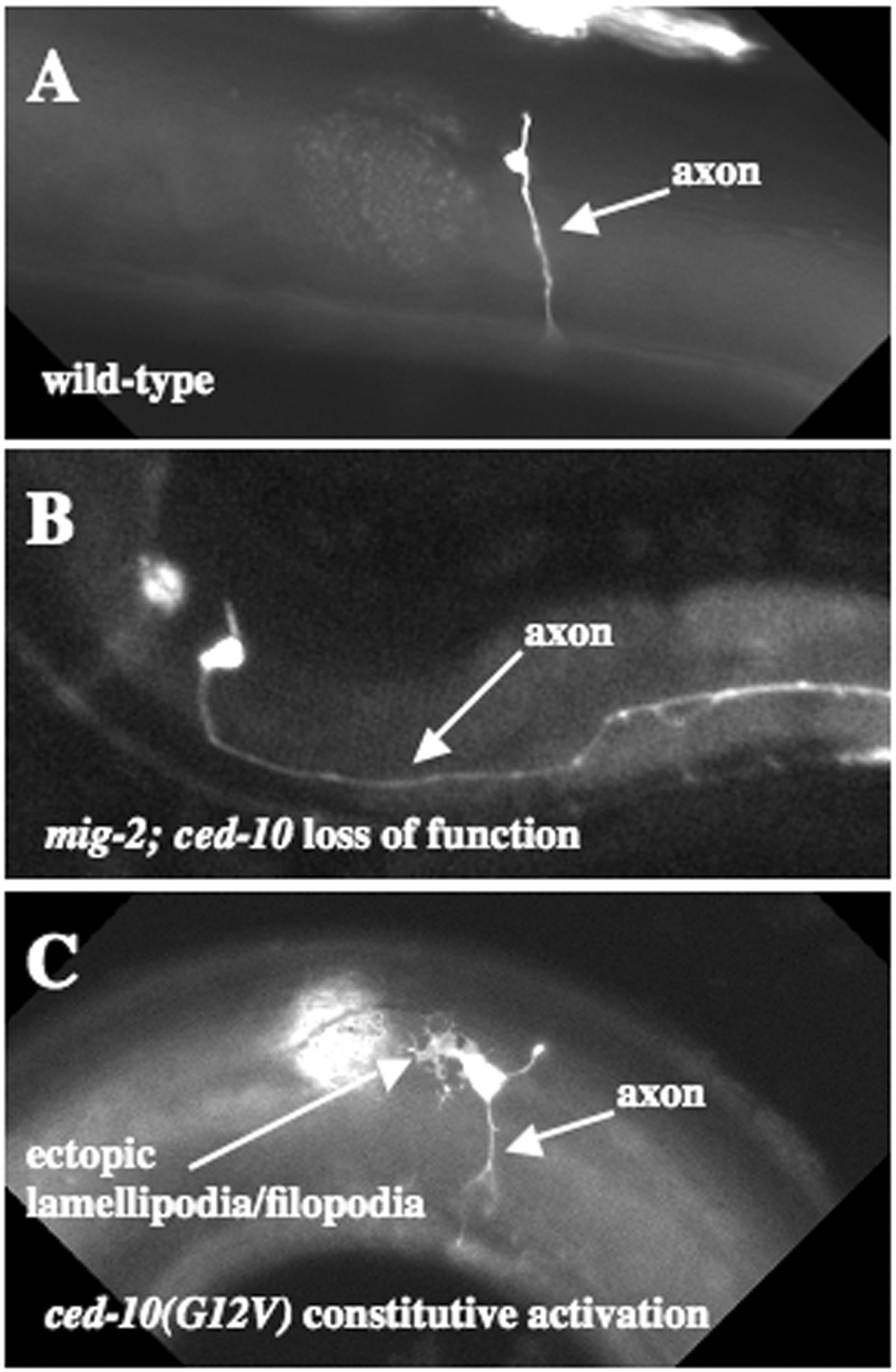
Null mutants of mig-2 and ced-10 and rac-2(RNAi) displayed no detectable defects in axon pathfinding (Lundquist et al., 2001; E.A.L., unpublished results). Each pairwise double mutant revealed synergistic axon pathfinding defects (i.e. the defects were more severe than the additive effects of each mutation alone; Lundquist et al., 2001; Wu et al., 2002; Figure 5). The double mutant phenotype included defects in axon outgrowth, axon guidance, and axon branching, indicating that Racs control multiple aspects of axon pathfinding. Furthermore, Rac double mutants displayed ectopic axon branches, suggesting that Racs also control the pruning of spurious axon branches (Struckhoff and Lundquist, 2003). Consistent with a role of Racs in axon development, constitutively-active CED-10, RAC-2 and MIG-2 perturb axon pathfinding (Struckhoff and Lundquist, 2003; Zipkin et al., 1997; Figure 5). Rac(G12V) molecules induce the formation of ectopic lamellipodia and filopodia on neurons that resemble those found on the growth cone during axon outgrowth (Knobel et al., 1999).
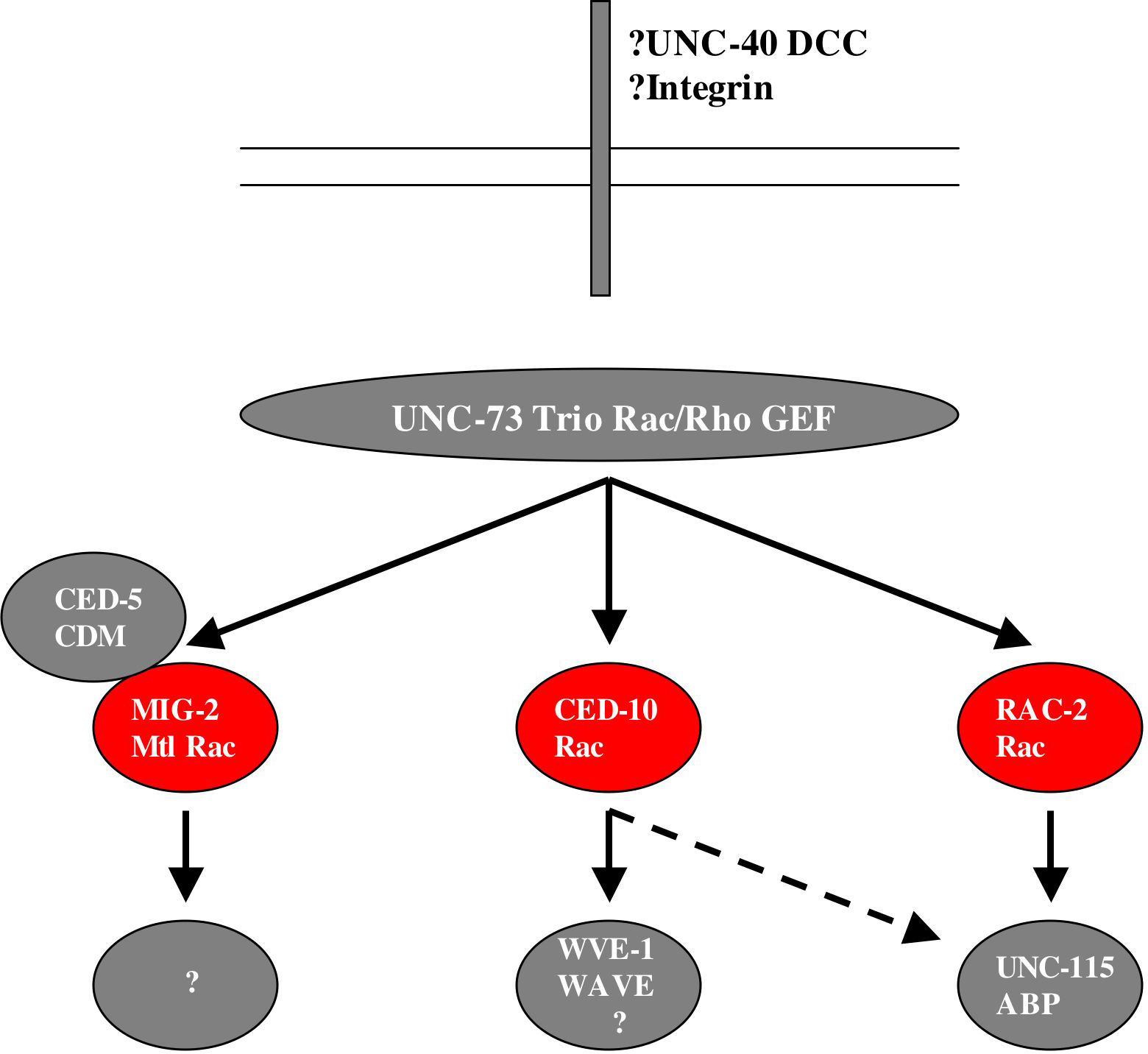
Using the logic that two mutations in the same redundant pathway should not enhance the other, whereas two mutations in distinct redundant pathways should show synergistic effects, other genes that act with rac signaling in axon pathfinding were identified (Figure 6). The UNC-73 Trio Rac/Rho GEF, which acts with RHO-1 in P cell migration, acts with all three Rac genes in axon pathfinding; CED-5, the CDM GEF that acts with in the CED-10 pathway in phagocytosis, acts in the MIG-2 pathway in parallel to CED-10 in axon pathfinding; and the actin-binding protein UNC-115 abLIM acts in the RAC-2 pathway in parallel to CED-10 and MIG-2 in axon pathfinding (Lundquist et al., 1998; Lundquist et al., 2001; Struckhoff and Lundquist, 2003; Wu et al., 2002). UNC-115 abLIM might represent one path to the actin cytoskeleton downstream of RAC-2. Indeed, unc-115 loss of fucntion partially suppresses the ectopic lamellipodia and filopodia induced by activated rac-2, indicating that unc-115 acts downstream of rac-2 in lamellipodia and filopodia formation (Struckhoff and Lundquist, 2003). RNAi of the Arp2/3 complex activator WVE-1 WAVE caused no axon defects alone but synergized with mig-2 mutations but not ced-10 mutations, suggesting that WVE-1 and possibly the Arp2/3 complex act downstream of CED-10 Rac in axon pathfinding (E.A.L., unpublished results). This combination of double mutant analysis and epistasis analysis will be important to identify additional genes that act downstream of Rac signaling in axon pathfinding and will contribute to a more complete understanding of the signaling mechanisms downstream of Rac.
Growth cones detect guidance information using transmembrane guidance receptors. Indeed, CED-10 Rac and the UNC-115 actin-binding protein act downstream of the netrin guidance receptor UNC-40 DCC in ventral AVM axon pathfinding (Gitai et al., 2003). Furthermore, CED-10, MIG-2 and RAC-2 act with the MIG-15 NIK kinase downstream of the integrin complex in VD/DD motor axon circumferential navigation (Poinat et al., 2002).
|
Figure 5. Axon pathfinding phenotypes of Rac mutants. Micrographs are lateral aspects of the posterior-lateral postdeirid region of adult C. elegans. Anterior is to the left and ventral is to the bottom. (A) A wild-type PDE neuron extends a single, unbranched axon ventrally to the ventral cord. (B) A mig-2; ced-10 double mutant PDE neuron displays a misguided axon that wanders laterally. (C) A PDE neuron with CED-10(G12V) expression exhibits ectopic neurites and ectopic lamellipodia and filopodia structures.
|
Figure 6. Overlapping roles of Racs in axon pathfinding. and neuronal migration. Three Rac proteins CED-10, MIG-2 and RAC-2 redundantly control axon pathfinding and neuonal migration. UNC-73 controls all three Racs in this process; the actin-binding protein UNC-115 acts downstream of RAC-2 and possibly CED-10; and the Arp2/3 activator WVE-1 WAVE acts in the CED-10 pathway. This network might act downstream of the UNC-6 netrin receptor UNC-40 DCC and integrin.
ced-10 and mig-2 mutations alone affect some cell migrations, including the gonadal distal tip cells and, in the case of mig-2, Q cells and their descendants. The migrations of the CAN neuron cell bodies, which are born in the anterior of the embryo and migrate posteriorly to a position near the vulva, are redundantly controlled by the Rac genes. No single Rac loss-of-function mutant displays CAN migration defects, but each pairwise double mutant combination of ced-10, mig-2 and rac-2 display synergistic failure in the posterior migrations of the CAN neurons (Lundquist et al., 2001).
Rac genes also act redundantly in multiple aspects of vulval development and morphogenesis. mig-2 and ced-10 single mutant vulvae are largely wild-type, whereas mig-2; ced-10 double loss-of-function mutants display defects in the orientiation of asymmetric divisions of the 1° and 2° vulval cells, indicating that mig-2 and ced-10 might redundantly control spindle orientation (Kishore and Sundaram, 2002). mig-2; ced-10 doubles also display a failure in the migrations of 2° vulval cells toward the 1° vulval cells to form a functional vulva. unc-73 mutations cause indistinguishable vulval defects, indicating that the UNC-73 Rho/Rac GEF controls both CED-10 and MIG-2 in vulval cell mitotic spindle orientation and 2° cell migration.
These studies of the roles of the three C. elegans Rac proteins have begun to elucidate the molecular complexes that participate with Racs in different developmental events (Figure 4). Further studies on Rac involvement in distinct cellular processes will decipher which Racs control which developmental events and what molecules act with the Racs in each process.
Rab GTPases are conserved regulators of multiple aspects of intracellular membrane trafficking and dynamics. In C. elegans there are 25 members of the Rab sub-family, and Rab genes are likely to have redundant roles in development as described for the Rho-family GTPases above. This might explain why traditional forward genetic screens have failed to identify mutations in most of the Rab genes. The RAB-1 and RAB-3 proteins have been identified in studies of innate immunity and synaptic vesicle trafficking, respectively.
Innate immunity in C. elegans consists of the expression and secretion of anti-microbial peptides, the neuropeptide-like proteins (NLPs) and the caenacins (CNCs; Couillault et al., 2004). Expression of nlp-31 in response to fungal infection requires the tir-1 gene, which encodes a Toll/IL/IR-domain protein similar to vertbrate SARM (Couillault et al., 2004). A yeast two-hybrid screen with TIR-1 identified RAB-1. RNAi of RAB-1 abolished nlp-31 expression in response to fungal infection (Couillault et al., 2004), indicating that RAB-1 mediates fungally-induced nlp-31 expression and might act with TIR-1 in this process. The molecular and cellular mechanism of RAB-1 in this process remains unclear.
The C. elegans RAB-3 protein is associated with acetylcholinergic synapses where it might influence synaptic vesicle trafficking and acetylcholine release (Nonet et al., 1997). Mutations in rab-3 were isolated in a screen for animals that were resistant to the acetylcholinesterase inhibitor aldicarb. Aldicarb causes overaccumulation of acetylcholine in the synaptic cleft which results in paralysis, and mutations in rab-3, which reduce the release of acetylcholine from presynaptic terminals, mitigate this effect. In rab-3 mutants, synaptic vesicles containing acetylcholine fail to be properly localized to acetylcholinergic synapses and accumulate in intersynaptic regions of the axon (Nonet et al., 1997). This phenotype is consistent with RAB-3 controlling the trafficking of synaptic vesicles to the synapse.
aex-3 encodes a Rab3 GEF, and mutations in aex-3 were isolated in a screen for defecation-defective animals (Iwasaki et al., 1997; Iwasaki and Toyonaga, 2000). In aex-3 mutants, RAB-3 protein accumulates in neuronal cell bodies and is not targeted to synapses in axons. Thus, AEX-3 might control the association of RAB-3 with synaptic vesicles.
In addition to affecting synaptic vesicle localization, RAB-3 might also control other aspects of vesicle release. unc-10 encodes a PDZ domain- and C2 domain-containing protein similar to the known Rab3 effector Rim1 (Koushika et al., 2001). Vesicle docking and synaptic organization appear normal in unc-10 mutants, and UNC-10 might interact with RAB-3 and syntaxin to facilitate vesicle fusion.
The single C. elegans Ran ortholog has been studied using RNAi. Consistent with studies in other systems in which Ran controls nuclear membrane organization and transport, ran-1(RNAi) embryos display failure of nuclear membrane reassembly after mitosis (Askjaer et al., 2002; Bamba et al., 2002). Additionally, ran-1(RNAi) causes defects in mitotic spindle formation consistent with a defect in kinetechore association with microtubules rather than a defect in microtubule organization per se. RNAi of genes associated with Ran signaling cause similar defects, including the GAP RanGAP, the Ran-binding protein RanBP2, and the Ran effector RCC1.
There are 12 annotated members of the Arf/Sar family of small GTPases in the C. elegans genome. Like the Rhos and the Rabs, the multiple Arf/Sar-family molecules might have overlapping roles in development and physiology and might have been missed in traditional forward-genetic screens. To date, two Arf/Sar-family molecule have been characterized in C. elegans.
evl-20 encodes an Arf protein of the Arl subtype, and mutations in the gene were identified in a forward genetic screen for animals with abnormal eversion of the vulva (the Evl phenotype; Antoshechkin and Han, 2002). In evl-20 mutants, the vulval precursor cells fail to produce the proper number of daughter cells, likely because of failed cytokinesis. As a result, vulval morphogenesis fails. EVL-20 is required for organization of the microtubule cytoskeleton, including the mitotic spindle involved in cleavage furrow positioning, and effects the morphogenesis of other tissues including the gonad and the hypodermis during embryonic development.
In C. elegans, ARL-6 is expressed in ciliated neurons and ARL-6::GFP fusion proteins localize to cilia (Fan et al., 2004). In fact, ARL-6::GFP appears to undergo intraflagellar transport and moves up and down the cilia possibly due to the action of +-end and –-end directed microtubule motors (kinesin and dynein, respectively). ARL-6 might be involved in ciliary development.
Studies of arl-6 in C. elegans represent a novel and powerful application of model organism genetics to human disease. Bardet-Beidl syndromes (BBSs) are multisystemic disorders of blindness, polydactyly, obesity, and cognitive impairment (Katsanis, 2004). Mutations at multiple loci can cause BBS, but the cellular and molecular mechanism of the disease is unclear. Interestingly, all BBS gene homologs in C. elegans are expressed in ciliated neurons (Li et al., 2004), indicating that the disease might affect ciliary function. This fact was used to identify the human gene responsible for BBS3 (Fan et al., 2004). C. elegans genes expressed in cilated neurons contain a DAF-19 RFX binding X-box in their promoters (Ansley et al., 2003; Li et al., 2004). All C. elegans genes with X-boxes in their promoters were identified, along with their counterparts in the human genome. Those human genes that localized to the genetically defined interval of BBS3 were sequenced in BBS3 patients. By this method, human arl-6 was found to harbor mutations in multiple BBS3 families (Fan et al., 2004).
Cellular and developmental roles for the 56 Ras-family small GTPases in the C. elegans genome are just beginning to be deciphered and include cell fate determination, gene expression, phagocytosis, cell migration and axon pathfinding, cytoskeletal organization, vesicle trafficking, and nuclear assembly (Table 2). Studies of small GTPases in C. elegans have been central to defining developmental roles for the Ras superfamily GTPases and in identifying and characterizing molecules that participate with GTPases in signaling networks. While much is known about let-60 Ras and Rho GTPase function, the roles of many members of the Ras/Ral/Rap, Rab, and Arf/Sar family remain to be discovered.
Table 2. Known roles of Ras-superfamily GTPases in C. elegans
| C. elegans gene name a | Human/mouse homolog b | Developmental role(s) c |
|---|---|---|
| Ras/Rap/Ral family | ||
| let-60 | K-Ras | Cell fate determination, oocyte meiotic maturation, sex myoblast migration, axon pathfinding, olfaction, response to microbacterium nematophilum infection |
| Rho family | ||
| rho-1 | RhoA | Cytokinesis, hypodermal contraction, P cell migration, axon pathfinding |
| cdc-42 | Cdc42 | Cytokinesis, embryonic polarity, axon pathfinding |
| ced-10 | Rac1 | Gastrulation, cell migration (P cells, distal tip cells), cell corpse phagocytosis, neuronal migration, axon pathfinding, vulval morphogenesis |
| rac-2 | Rac1 | Neuronal migration, axon pathfinding |
| mig-2 | Mtl | Cell migration (P-cells, distal tip cells, Q cells and descendants), neuronal migration, axon pathfinding, vulval morphogenesis |
| Rab family | ||
| rab-1 | Rab1 | Innate immunity gene expression |
| rab-3 | Rab3 | Synaptic vesicle trafficking and release |
| Arf/Sar family | ||
| arl-6 | Arf6 | Ciliary vesicle trafficking (?) |
| evl-20 | Arf2 | Microtubule organization, vulval cell division, tissue morphogenesis |
| Ran family | ||
| ran-1 | Ran | Nuclear trafficking, nuclear reassembly, Kinetechore association with the mitotic spindle |
|
aThe three-letter C. elegans gene designation. Not all genes have a three letter designation. bSimilarities are as noted in Wormbase. cSee text for details. |
||
Studies of Ras-superfamily GTPases in C. elegans have demonstrated that many of the GTPases have overlapping, redundant roles in developmental processes, including Rap GTPases in hypodermal morphogenesis and function and Rho GTPases in cell migration and axon pathfinding. The multiple Rab and Arf/Sar GTPases might display similar redundancy of function, explaining why they have not been identified in forward genetic screens. These studies also demonstrate that the same GTPase can act with different signaling partners in different developmental events. For example, the UNC-73 Trio Rac/Rho GEF acts with CED-10 Rac in axon pathfinding but not in phagocytosis (Figure 4). Further genetic and molecular analyses will be required to decipher the roles of all Ras-superfamily GTPases in development and to delineate the signaling pathways by which they control different developmental events.
I thank members of the Lundquist lab for helpful comments and discussion. Rho GTPase work in my lab is supported by NIH grant NS40945 and NSF grant IBN93192.
Ansley, S.J., Badano, J.L., Blacque, O.E., Hill, J., Hoskins, B.E., Leitch, C.C., Kim, J.C., Ross, A.J., Eichers, E.R., Teslovich, T.M., et al. (2003).Basal body dysfunction is a likely cause of pleiotropic Bardet-Biedl syndrome. Nature 425,628–633. Abstract Article
Antoshechkin, I., and Han, M. (2002). The C. elegans evl-20 gene is a homolog of the small GTPase ARL2 and regulates cytoskeleton dynamics during cytokinesis and morphogenesis. Dev. Cell 2, 579–591. Abstract Article
Askjaer, P., Galy, V., Hannak, E., and Mattaj, I.W. (2002). Ran GTPase cycle and importins α and Β are essential for spindle formation and nuclear envelope assembly in living Caenorhabditis elegans embryos. Mol. Biol. Cell 13, 4355–4370. Abstract Article
Bamba, C., Bobinnec, Y., Fukuda, M., and Nishida, E. (2002). The GTPase Ran regulates chromosome positioning and nuclear envelope assembly in vivo. Curr. Biol. 12, 503–507. Abstract Article
Beitel, G.J., Clark, S.G., and Horvitz, H.R. (1990).Caenorhabditis elegans ras gene let-60 acts as a switch in the pathway of vulval induction. Nature 348, 503–509. Abstract Article
Berkel, W.P., Verheijen, M.H., Cuppen, E., Asahina, M., de Rooij, J., Jansen, G., Plasterk, R.H., Bos, J.L., and Zwartkruis, F.J. (2005). Requirement of the Caenorhabditis elegans RapGEF pxf-1 and rap-1 for Epithelial Integrity. Mol. Biol. Cell 16, 106–116. Abstract Article
Bos, J.L., and Zwartkruis, F.J. (1999). Signal transduction. Rhapsody in G proteins. Nature 400, 820–821. Abstract Article
Bourne, H.R., Sanders, D.A., and McCormick, F. (1991). The GTPase superfamily: conserved structure and molecular mechanism. Nature 349, 117–127. Abstract Article
Buechner, M., Hall, D.H., Bhatt, H., and Hedgecock, E.M. (1999). Cystic canal mutants in Caenorhabditis elegans are defective in the apical membrane domain of the renal (excretory) cell. Dev. Biol. 214, 227–241. Abstract Article
Church, D.L., Guan, K.L., and Lambie, E.J. (1995). Three genes of the MAP kinase cascade, mek-2, mpk-1/sur-1 and let-60 ras, are required for meiotic cell cycle progression in Caenorhabditis elegans. Development 121, 2525–2535. Abstract
Clark, S.G., Stern, M.J., and Horvitz, H.R. (1992). C. elegans cell-signalling gene sem-5 encodes a protein with SH2 and SH3 domains. Nature 356, 340–344. Abstract Article
Couillault, C., Pujol, N., Reboul, J., Sabatier, L., Guichou, J.F., Kohara, Y., and Ewbank, J.J. (2004). TLR-independent control of innate immunity in Caenorhabditis elegans by the TIR domain adaptor protein TIR-1, an ortholog of human SARM. Nat. Immunol. 5, 488–494. Abstract Article
deBakker, C.D., Haney, L.B., Kinchen, J.M., Grimsley, C., Lu, M., Klingele, D., Hsu, P.K., Chou, B.K., Cheng, L.C., Blangy, A., et al. (2004). Phagocytosis of apoptotic cells is regulated by a UNC-73/TRIO-MIG-2/RhoG signaling module and armadillo repeats of CED-12/ELMO. Curr. Biol. 14, 2208–2216. Abstract Article
Eden, S., Rohatgi, R., Podtelejnikov, A.V., Mann, M., and Kirschner, M.W. (2002). Mechanism of regulation of WAVE1-induced actin nucleation by Rac1 and Nck. Nature 418, 790–793. Abstract Article
Ellis, R.E., Jacobson, D.M., and Horvitz, H.R. (1991). Genes required for the engulfment of cell corpses during programmed cell death in Caenorhabditis elegans. Genetics 129, 79–94. Abstract
Etemad-Moghadam, B., Guo, S., and Kemphues, K.J. (1995). Asymmetrically distributed PAR-3 protein contributes to cell polarity and spindle alignment in early C. elegans embryos. Cell 83, 743–752. Abstract Article
Fan, Y., Esmail, M.A., Ansley, S.J., Blacque, O.E., Boroevich, K., Ross, A. J., Moore, S.J., Badano, J.L., May-Simera, H., Compton, D.S., et al. (2004). Mutations in a member of the Ras superfamily of small GTP-binding proteins causes Bardet-Biedl syndrome. Nat. Genet. 36, 989–993. Abstract Article
Ferguson, E.L., and Horvitz, H.R. (1985). Identification and characterization of 22 genes that affect the vulval cell lineages of the nematode Caenorhabditis elegans. Genetics 110, 17–72. Abstract
Ferguson, E.L., Sternberg, P.W., and Horvitz, H.R. (1987). A genetic pathway for the specification of the vulval cell lineages of Caenorhabditis elegans. Nature 326, 259–267. Abstract Article
Gao, J., Estrada, L., Cho, S., Ellis, R.E., and Gorski, J.L. (2001). The Caenorhabditis elegans homolog of FGD1, the human Cdc42 GEF gene responsible for faciogenital dysplasia, is critical for excretory cell morphogenesis. Hum. Mol. Genet. 10, 3049–3062. Abstract Article
Gitai, Z., Yu, T.W., Lundquist, E.A., Tessier-Lavigne, M., and Bargmann, C.I. (2003). The netrin receptor UNC-40/DCC stimulates axon attraction and outgrowth through Enabled and, in parallel, Rac and UNC-115/AbLIM. Neuron 37, 53–65. Abstract Article
Gotta, M., Abraham, M.C., and Ahringer, J. (2001). CDC-42 controls early cell polarity and spindle orientation in C. elegans. Curr. Biol. 11, 482–488. Abstract Article
Gumienny, T.L., Brugnera, E., Tosello-Trampont, A.C., Kinchen, J.M., Haney, L.B., Nishiwaki, K., Walk, S.F., Nemergut, M.E., Macara, I.G., Francis, R., et al. (2001). CED-12/ELMO, a novel member of the CrkII/Dock180/Rac pathway, is required for phagocytosis and cell migration. Cell 107, 27–41. Abstract Article
Han, M., Aroian, R.V., and Sternberg, P.W. (1990). The let-60 locus controls the switch between vulval and nonvulval cell fates in Caenorhabditis elegans. Genetics 126, 899–913. Abstract
Han, M., and Sternberg, P.W. (1990). let-60, a gene that specifies cell fates during C. elegans vulval induction, encodes a ras protein. Cell 63, 921–931. Abstract Article
Hung, T.J., and Kemphues, K.J. (1999). PAR-6 is a conserved PDZ domain-containing protein that colocalizes with PAR-3 in Caenorhabditis elegans embryos. Development 126, 127–135. Abstract
Iwasaki, K., Staunton, J., Saifee, O., Nonet, M., and Thomas, J.H. (1997). aex-3 encodes a novel regulator of presynaptic activity in C. elegans. Neuron 18, 613–622. Abstract Article
Iwasaki, K., and Toyonaga, R. (2000). The Rab3 GDP/GTP exchange factor homolog AEX-3 has a dual function in synaptic transmission. EMBO J. 19, 4806–4816. Abstract Article
Jantsch-Plunger, V., Gonczy, P., Romano, A., Schnabel, H., Hamill, D., Schnabel, R., Hyman, A.A., and Glotzer, M. (2000). CYK-4: a Rho family gtpase activating protein (GAP) required for central spindle formation and cytokinesis. J. Cell Biol. 149, 1391–1404. Abstract Article
Katsanis, N. (2004). The oligogenic properties of Bardet-Biedl syndrome. Hum. Mol. Genet. 13(Spec No 1), R65–R71. Abstract Article
Kay, A.J., and Hunter, C.P. (2001). CDC-42 regulates PAR protein localization and function to control cellular and embryonic polarity in C. elegans. Curr. Biol. 11, 474–481. Abstract Article
Kemphues, K.J., Priess, J.R., Morton, D.G., and Cheng, N.S. (1988). Identification of genes required for cytoplasmic localization in early C. elegans embryos. Cell 52, 311–320. Abstract Article
Kishore, R.S., and Sundaram, M.V. (2002). ced-10 Rac and mig-2 function redundantly and act with unc-73 trio to control the orientation of vulval cell divisions and migrations in Caenorhabditis elegans. Dev. Biol. 241, 339–348. Abstract Article
Knobel, K.M., Jorgensen, E.M., and Bastiani, M.J. (1999). Growth cones stall and collapse during axon outgrowth in Caenorhabditis elegans. Development 126, 4489–4498. Abstract
Koushika, S.P., Richmond, J.E., Hadwiger, G., Weimer, R.M., Jorgensen, E.M., and Nonet, M.L. (2001). A post-docking role for active zone protein Rim. Nat. Neurosci. 4, 997–1005. Abstract Article
Levitan, D.J., Boyd, L., Mello, C.C., Kemphues, K.J., and Stinchcomb, D.T. (1994). par-2, a gene required for blastomere asymmetry in Caenorhabditis elegans, encodes zinc-finger and ATP-binding motifs. Proc. Natl. Acad. Sci. USA 91, 6108–6112. Abstract
Li, J.B., Gerdes, J.M., Haycraft, C.J., Fan, Y., Teslovich, T.M., May-Simera, H., Li, H., Blacque, O.E., Li, L., Leitch, C.C., et al. (2004). Comparative genomics identifies a flagellar and basal body proteome that includes the BBS5 human disease gene. Cell 117, 541–552. Abstract Article
Lu, M., Kinchen, J.M., Rossman, K.L., Grimsley, C., deBakker, C., Brugnera, E., Tosello-Trampont, A.C., Haney, L.B., Klingele, D., Sondek, J., et al. (2004). PH domain of ELMO functions in trans to regulate Rac activation via Dock180. Nat. Struct. Mol. Biol. 11, 756–762. Abstract Article
Lundquist, E.A., Herman, R.K., Shaw, J.E., and Bargmann, C.I. (1998). UNC-115, a conserved protein with predicted LIM and actin-binding domains, mediates axon guidance in C. elegans. Neuron 21, 385–392. Abstract Article
Lundquist, E.A., Reddien, P.W., Hartwieg, E., Horvitz, H.R., and Bargmann, C.I. (2001). Three C. elegans Rac proteins and several alternative Rac regulators control axon guidance, cell migration and apoptotic cell phagocytosis. Development 128, 4475–4488. Abstract
Nonet, M.L., Staunton, J.E., Kilgard, M.P., Fergestad, T., Hartwieg, E., Horvitz, H.R., Jorgensen, E.M., and Meyer, B.J. (1997). Caenorhabditis elegans rab-3 mutant synapses exhibit impaired function and are partially depleted of vesicles. J. Neurosci. 17, 8061–8073. Abstract
Piekny, A.J., and Mains, P.E. (2002). Rho-binding kinase (LET-502) and myosin phosphatase (MEL-11) regulate cytokinesis in the early Caenorhabditis elegans embryo. J. Cell Sci. 115, 2271–2282. Abstract
Piekny, A.J., Wissmann, A., and Mains, P.E. (2000). Embryonic morphogenesis in Caenorhabditis elegans integrates the activity of LET-502 Rho-binding kinase, MEL-11 myosin phosphatase, DAF-2 insulin receptor and FEM-2 PP2c phosphatase. Genetics 156, 1671–1689. Abstract
Poinat, P., De Arcangelis, A., Sookhareea, S., Zhu, X., Hedgecock, E.M., Labouesse, M., and Georges-Labouesse, E. (2002). A conserved interaction between Β1 integrin/PAT-3 and Nck-interacting kinase/MIG-15 that mediates commissural axon navigation in C. elegans. Curr. Biol. 12, 622–631. Abstract Article
Price, L.S., Hajdo-Milasinovic, A., Zhao, J., Zwartkruis, F.J., Collard, J.G., and Bos, J.L. (2004). Rap1 regulates E-cadherin-mediated cell-cell adhesion. J. Biol. Chem. 279, 35127–35132. Abstract Article
Rangarajan, S., Enserink, J.M., Kuiperij, H.B., de Rooij, J., Price, L.S., Schwede, F., and Bos, J.L. (2003). Cyclic AMP induces integrin-mediated cell adhesion through Epac and Rap1 upon stimulation of the Β 2-adrenergic receptor. J. Cell Biol. 160, 487–493. Abstract Article
Reddien, P.W., and Horvitz, H.R. (2000). CED-2/CrkII and CED-10/Rac control phagocytosis and cell migration in Caenorhabditis elegans. Nat. Cell Biol. 2, 131–136. Abstract Article
Simske, J.S., and Hardin, J. (2001). Getting into shape: epidermal morphogenesis in Caenorhabditis elegans embryos. Bioessays 23, 12–23. Abstract
Soto, M.C., Qadota, H., Kasuya, K., Inoue, M., Tsuboi, D., Mello, C.C., and Kaibuchi, K. (2002). The GEX-2 and GEX-3 proteins are required for tissue morphogenesis and cell migrations in C. elegans. Genes Dev. 16, 620–632. Abstract Article
Spencer, A.G., Orita, S., Malone, C.J., and Han, M. (2001). A RHO GTPase-mediated pathway is required during P cell migration in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 98, 13132–13137. Abstract Article
Stern, M.J., Marengere, L.E., Daly, R.J., Lowenstein, E.J., Kokel, M., Batzer, A., Olivier, P., Pawson, T., and Schlessinger, J. (1993). The human GRB2 and Drosophila Drk genes can functionally replace the Caenorhabditis elegans cell signaling gene sem-5. Mol. Biol. Cell 4, 1175–1188. Abstract
Steven, R., Kubiseski, T.J., Zheng, H., Kulkarni, S., Mancillas, J., Ruiz Morales, A., Hogue, C.W., Pawson, T., and Culotti, J. (1998). UNC-73 activates the Rac GTPase and is required for cell and growth cone migrations in C. elegans. Cell 92, 785–795. Abstract Article
Struckhoff, E.C., and Lundquist, E.A. (2003). The actin-binding protein UNC-115 is an effector of Rac signaling during axon pathfinding in C. elegans. Development 130, 693–704. Abstract Article
Sulston, J.E., Schierenberg, E., White, J.G., and Thomson, J.N. (1983). The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100, 64–119. Abstract Article
Sundaram, M., Yochem, J., and Han, M. (1996). A Ras-mediated signal transduction pathway is involved in the control of sex myoblast migration in Caenorhabditis elegans. Development 122, 2823–2833. Abstract
Suzuki, N., Buechner, M., Nishiwaki, K., Hall, D.H., Nakanishi, H., Takai, Y., Hisamoto, N., and Matsumoto, K. (2001). A putative GDP-GTP exchange factor is required for development of the excretory cell in Caenorhabditis elegans. EMBO Rep. 2, 530–535. Abstract Article
Tao, W., Pennica, D., Xu, L., Kalejta, R.F., and Levine, A.J. (2001). Wrch-1, a novel member of the Rho gene family that is regulated by Wnt-1. Genes Dev. 15, 1796–1807. Abstract Article
Watts, J.L., Etemad-Moghadam, B., Guo, S., Boyd, L., Draper, B.W., Mello, C.C., Priess, J.R., and Kemphues, K.J. (1996). par-6, a gene involved in the establishment of asymmetry in early C. elegans embryos, mediates the asymmetric localization of PAR-3. Development 122, 3133–3140. Abstract
Williams-Masson, E.M., Malik, A.N., and Hardin, J. (1997). An actin-mediated two-step mechanism is required for ventral enclosure of the C. elegans hypodermis. Development 124, 2889–2901. Abstract
Wissmann, A., Ingles, J., and Mains, P.E. (1999). The Caenorhabditis elegans mel-11 myosin phosphatase regulatory subunit affects tissue contraction in the somatic gonad and the embryonic epidermis and genetically interacts with the Rac signaling pathway. Dev. Biol. 209, 111–127. Abstract Article
Wissmann, A., Ingles, J., McGhee, J.D., and Mains, P.E. (1997). Caenorhabditis elegans LET-502 is related to Rho-binding kinases and human myotonic dystrophy kinase and interacts genetically with a homolog of the regulatory subunit of smooth muscle myosin phosphatase to affect cell shape. Genes Dev. 11, 409–422. Abstract
Withee, J., Galligan, B., Hawkins, N., and Garriga, G. (2004). Caenorhabditis elegans WASP and Ena/VASP proteins play compensatory roles in morphogenesis and neuronal cell migration. Genetics 167, 1165–1176. Abstract Article
Wu, Y.C., Cheng, T.W., Lee, M.C., and Weng, N.Y. (2002). Distinct rac activation pathways control Caenorhabditis elegans cell migration and axon outgrowth. Dev. Biol. 250, 145–155. Abstract Article
Wu, Y.C., and Horvitz, H.R. (1998). C. elegans phagocytosis and cell-migration protein CED-5 is similar to human DOCK180. Nature 392, 501–504. Abstract Article
Zallen, J.A., Peckol, E.L., Tobin, D.M., and Bargmann, C.I. (2000). Neuronal cell shape and neurite initiation are regulated by the Ndr kinase SAX-1, a member of the Orb6/COT-1/warts serine/threonine kinase family. Mol. Biol. Cell 11, 3177–3190. Abstract
*Edited by Iva Greenwald. Last revised January 20, 2005. Published January 17, 2006. This chapter should be cited as: Lundquist, E. A. Small GTPases (January 17, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.67.1, http://www.wormbook.org.
Copyright: © 2006 Erik A. Lundquist. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: erikl@ku.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.