Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Spermatogenesis creates functional sperm from an initially undifferentiated germ cell. In the nematode Caenorhabditis elegans, both males and hermaphrodites engage in spermatogenesis. The hermaphrodite germ line, like that of the male, initiates spermatogenesis during the L4 larval stage. The hermaphrodite germ line differs from that of the male because it ceases spermatogenesis and switches to oogenesis during the adult stage. Each hermaphrodite stores her sperm and uses them to fertilize her oocytes. Many mutants have been identified where hermaphrodite self-fertility is disrupted. If such a self-sterile hermaphrodite is mated to a wild-type male, mutant hermaphrodites that either lack sperm or contain defective sperm will produce outcross progeny. Easily implemented tests are then applied to identify the subset of these mutants that produce defective sperm. Currently, more than 44 genes are known that are required for normal spermatogenesis. This chapter discusses the 25 best-understood genes that affect spermatogenesis and mutants are grouped based on the cellular structure or process that is affected. C. elegans spermatozoa lack an acrosome and a flagellum, which are organelles found in the spermatozoa produced by most other species. Like other nematodes, C. elegans spermatozoa move by crawling using a single pseudopod. Wild-type spermatogenesis and its defects in mutants can be studied in vivo because the animal is transparent and in vitro because a simple, chemically defined medium that supports development has been discovered. Unlike nearly all other C. elegans cells, homogeneous sperm can be obtained in sufficient quantities to permit biochemical analyses.
Spermatogenesis is the process that creates sperm from initially undifferentiated germ cells (see Sex determination in the germ line). In diocious animals, this process occurs exclusively in males. While C. elegans spermatogenesis occurs in males, it also occurs during a stage of germline development in the hermaphrodite prior to the onset of oogenesis (see Sex determination in the germ line). Wild-type spermatogenesis and its defects in mutants can be studied in vivo because the animal is transparent and in vitro because a medium that supports development is available (Machaca et al., 1996; Nelson and Ward, 1980). Unlike nearly all other C. elegans cells, homogeneous sperm can be obtained in sufficient quantities to permit biochemical analyses (Klass and Hirsh, 1981; Nelson et al., 1982). This chapter focuses on genes that are sperm-specific regulators, and genes affecting germline development more generally are described elsewhere (see Sex determination in the germ line, RNA-binding proteins, Germline proliferation and its control and Specification of the germ line).
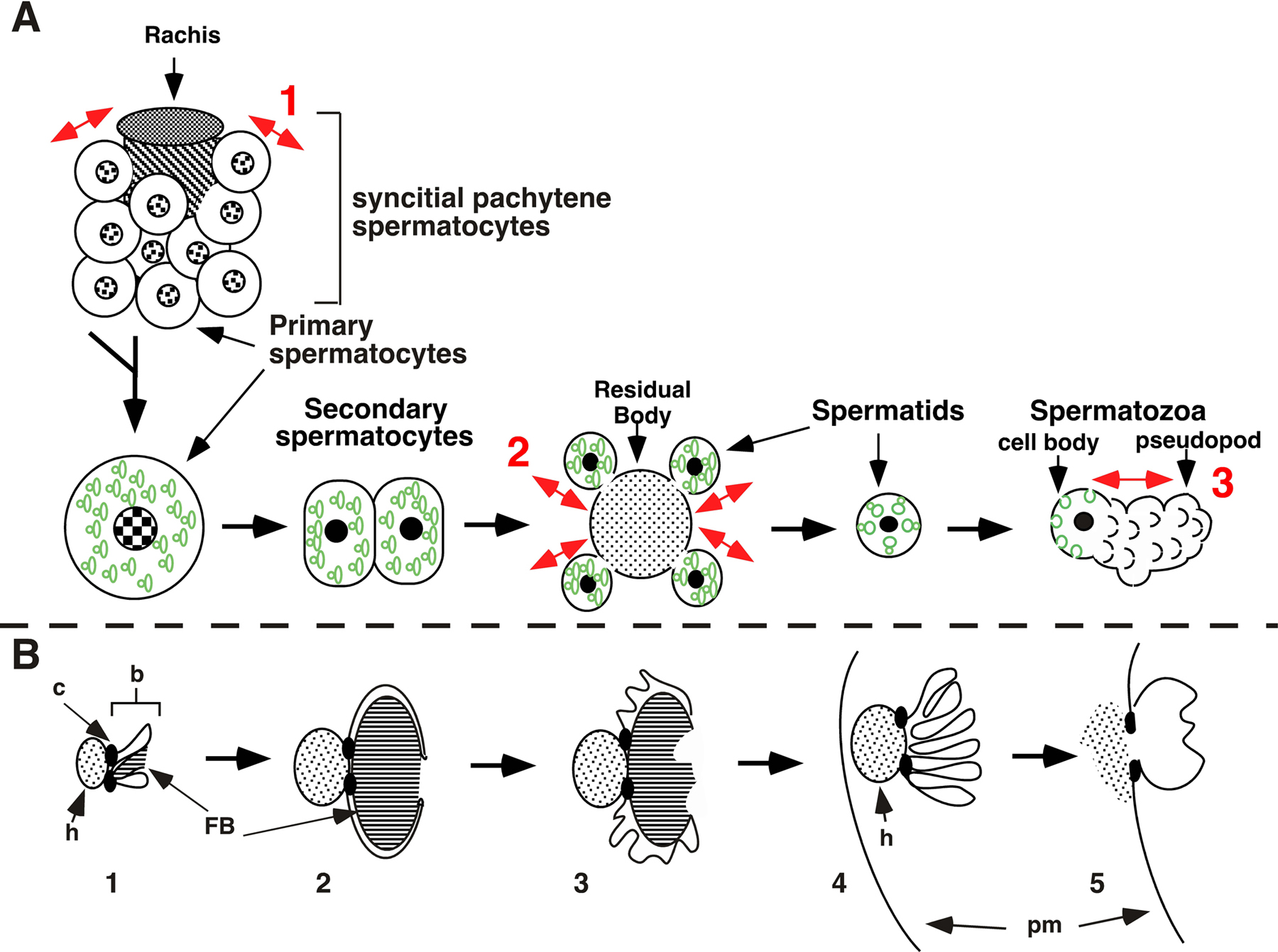
Development of sperm in C. elegans males has been described in detail (Kimble and Ward, 1988; Ward, 1986; Ward et al., 1981; Wolf et al., 1978) and these cellular events are reviewed in Figure 1. The primary spermatocyte initially forms in syncytium with a cytoplasmic core called the rachis. Upon entering meiosis, the primary spermatocyte buds off the rachis and completes development without any requirement for intimate association with other cells. As in most cells, nuclear divisions are closely coordinated with cytokinesis and other aspects of cytodifferentiation. Meiosis I divides the 4N primary spermatocyte nucleus into two 2N nuclei in secondary spermatocytes. The cytokinesis that accompanies meiosis I is either complete (not shown) or partial (2 in Figure 1A). Either way, the resulting cells are secondary spermatocytes and they immediately initiate meiosis II. Meiosis II produces two haploid spermatids from each 2N secondary spermatocyte. Spermatids form by budding from an anucleate residual body (Movie 1), and spermatids lack many cellular constituents present in the secondary spermatocyte. Material within the residual body, which includes most voltage-gated ion channels, tubulin, actin and all ribosomes (Machaca et al., 1996; Ward, 1986) is apparently resorbed (Kimble and Ward, 1988; Nelson et al., 1982; Ward, 1986; Ward et al., 1981; Wolf et al., 1978). Sessile spermatids lack ribosomes and have no new protein synthesis as they complete differentiation into motile spermatozoa (Ward et al., 1983). The cytologically obvious structures that segregate into spermatids during budding include its nucleus, multiple mitochondria and multiple specialized lysosome-like organelles named fibrous body-membranous organelles (FB-MOs).
The meiotic divisions of spermatogenesis are coordinated with the morphogenesis of FB-MOs (Ward et al., 1981; Wolf et al., 1978). In syncytial pachytene spermatocytes (Figure 1), the MOs form from the Golgi apparatus and FBs form in close association with the MOs (Roberts et al., 1986). Growth of the FBs occurs within a MO-derived membrane envelope in primary spermatocytes (Figure 1B1). As spermatids bud, the double membrane surrounding the FB retracts into the MO and the filamentous major sperm protein (MSP) contents (Figure 1B3) depolymerize and disperse throughout the spermatid cytoplasm. These FB-free MOs (Figure 1B4) localize near the spermatid plasma membrane (pm in Figure 1B4). Spermiogenesis is the process during which a spermatid becomes a spermatozoon. MO head membranes (h, in Figure 1B4) fuse and deposit their contents on the plasma membrane of the cell body as the spermatozoon forms (Figure 1B5). A single pseudopod is extended (red arrows at 3 in Figure 1A), and MSP fibers are confined to the pseudopod (Ward and Klass, 1982) while the nucleus, mitochondria, MOs and other internal membranes are confined to the cell body. Like other nematodes, C. elegans spermatozoa lack a flagellum and crawl using this single pseudopod (see Sperm motility and MSP).
|
Figure 1. (A). Summary of spermatogenesis. Asymmetric partitioning of cellular constituents occurs at three points during C. elegans spermatogenesis, as indicated by double-headed red arrows at the red numbers. 1, syncytial pachytene spermatocytes with many FB-MOs bud from the rachis and divide to form secondary spermatocytes (FB-MOs are shown in green); 2, spermatids selectively retain FB-MOs as they bud from the residual body; 3, FB-MOs fuse with the spermatid plasma membrane as a pseudopod extends from the cell body during spermiogenesis. Nuclei are the circles in the center of each cell. Nuclei are patterned with lines to represent stages when chromatin is in condensed meiotic chromosomes or filled (in black) after chromatin forms a single highly condensed sphere. (B) Summary of morphogenesis of the FB-MO complex. 1, The fibrous body (FB) develops in close association with, and is surrounded by, the membranous organelle (MO) within the primary spermatocyte. The MO is separated by a collar region (c) into a head (h; speckled region at left) and body (b; region to the right of the collar); 2, the FB-MO complex reaches its largest size within primary spermatocytes. The double layered MO-derived membrane surrounds the FB, which is composed of the major sperm protein filaments; 3, the MO-derived membranes surrounding the FB retract and fold up while FB filaments depolymerize and disperse as spermatids bud from the residual body; 4, the head of each MO (arrow) moves to a position just below the plasma membrane (pm) of the spermatid after the FBs have depolymerized and disappeared. The irregular shapes within the FB represent retracted membrane that had covered the exterior of the FB; 5, the head of the MO fuses at the collar to the plasma membrane and exocytoses its contents (dots at arrow) onto the cell surface. A permanent fusion pore remains at the point of each MO fusion (each cell has many MO's; B is modified from L'Hernault and Arduengo, 1992).
Figure 2. Video micrograph of spermatid budding. Two connected secondary spermatocytes form four spermatids and place much of the cytoplasm in the centrally located residual body. QuickTime movie courtesy of P. J. Muhlrad and S. Ward, University of Arizona.
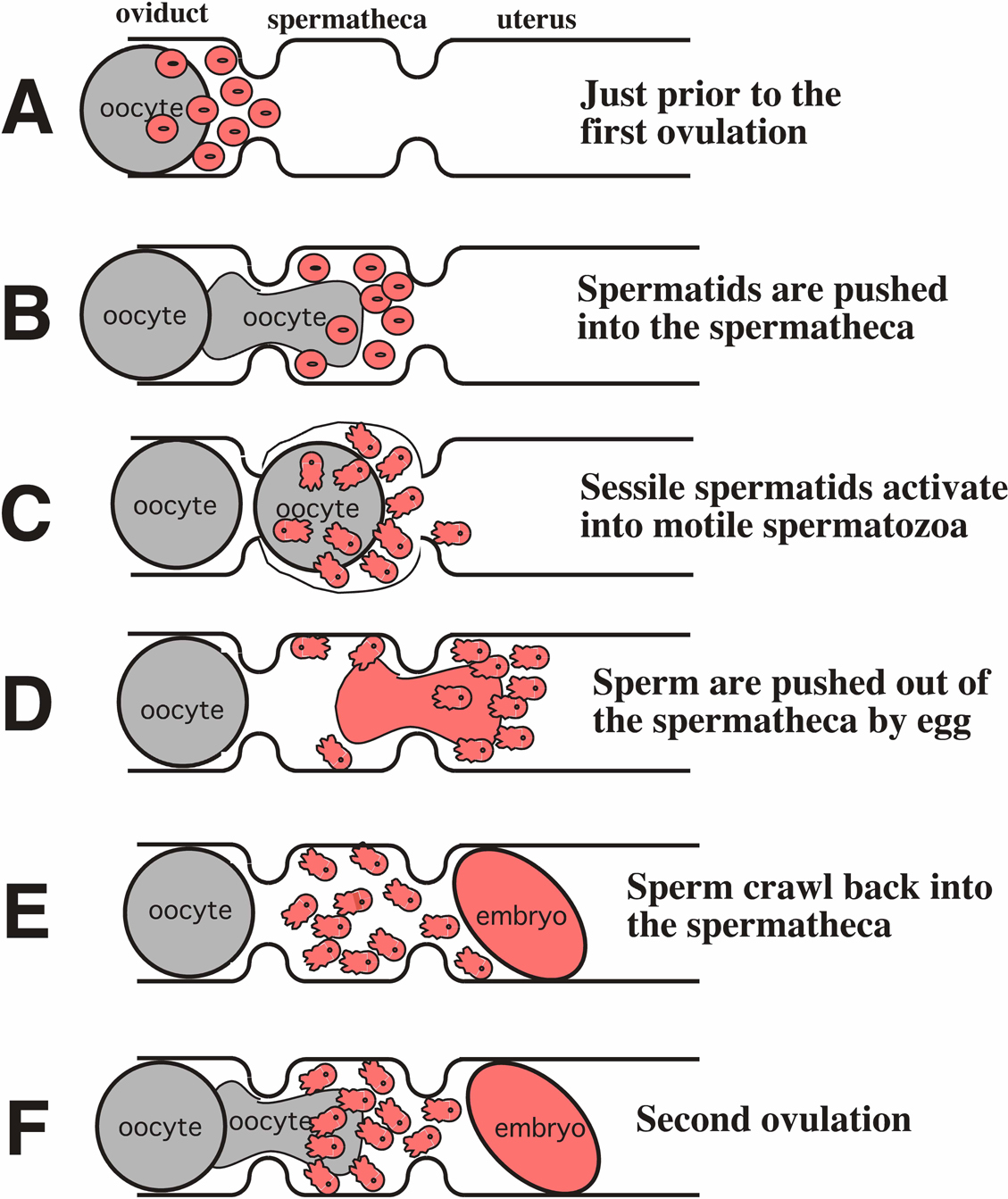
Spermatogenesis is similar in hermaphrodites and males, but there are some significant differences. Spermatids accumulate in the proximal gonad arm after the hermaphrodite germ line has switched to oogenesis (Figure 3A). The first ovulation pushes spermatids into the spermatheca (Figure 3B), where they rapidly undergo spermiogenesis and become spermatozoa (Figure 3C). Many spermatozoa are displaced into the uterus as the egg leaves the spermatheca and they must crawl back into the spermatheca in order to compete for oocytes (Figure 3D-F; Ward and Carrel, 1979).
|
Figure 3. Summary of gamete interactions in the hermaphrodite during self-fertilization. (A) Spermatids (in pink) are crowded at the oviduct spermathecal junction by maturing oocytes. (B) Spermatids are pushed into the spermatheca by the first ovulation. (C) Spermatids undergo spermiogenesis to become spermatozoa and compete with each other to fertilize the oocyte. (D) The newly fertilized egg exits the spermatheca and pushes many sperm into the uterus. (E) The displaced sperm crawl from the uterus back into the spermatheca. (F) The second ovulation places an oocyte in the spermatheca and sperm compete to fertilize it. The cell body is pulled behind the pseudopod as spermatozoa crawl forward.
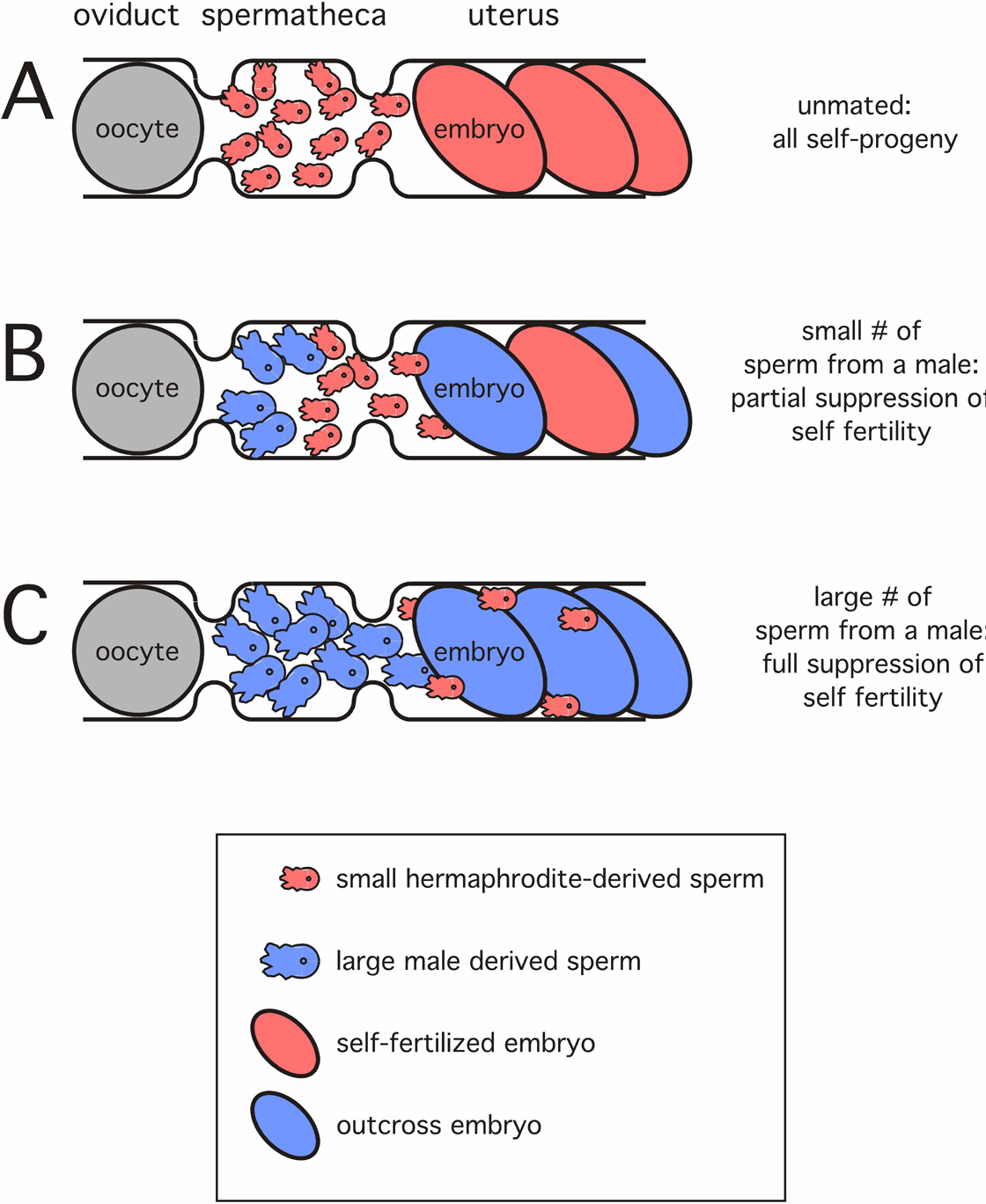
In males, spermatids are stored in the seminal vesicle. These spermatids are significantly larger than those produced by hermaphrodites (LaMunyon and Ward, 1998). Spermatids undergo spermiogenesis after they are mixed with seminal fluid during ejaculation into the hermaphrodite uterus. The molecular mechanism that activates spermiogenesis in vivo is not known for either sex. However, in vitro, spermatids are activated to become spermatozoa by treatment with several unrelated agents including the ionophore monensin (Nelson and Ward, 1980), proteases, weak bases that alkalinize the cytoplasm (Ward et al., 1983), calmodulin inhibitors (Shakes and Ward, 1989) or the ion channel inhibitor DIDS (Machaca et al., 1996). Normally, male-derived spermatozoa crawl towards and into the hermaphrodite spermatheca. Spermatozoa introduced by a male during copulation have a competitive advantage over hermaphrodite-derived spermatozoa in the struggle to fertilize oocytes (Figure 4; Ward and Carrel, 1979; LaMunyon and Ward, 1995). Artificial insemination is also possible, and male-derived spermatids that are protease treated cannot fertilize oocytes, while spermatids treated with weak bases are competent to participate in fertilization (LaMunyon and Ward, 1994).
|
Figure 4. Sperm competition. (A) Hermaphrodite-derived spermatozoa wait in the spermatheca, where they fertilize nearly all oocytes that pass through the spermatheca. (B) A hermaphrodite after being inseminated by relatively few male-derived (blue) sperm will produce a mixture of self (pink embryo) and outcross (blue embryo) progeny. (C) A hermaphrodite after being inseminated by many male-derived (blue) sperm will produce only outcross (blue embryo) progeny. As noted by others, male-derived sperm are ~50% larger than hermaphrodite-derived sperm (LaMunyon and Ward, 1995; LaMunyon and Ward, 1998). Adapted from Singson et al. (1999).
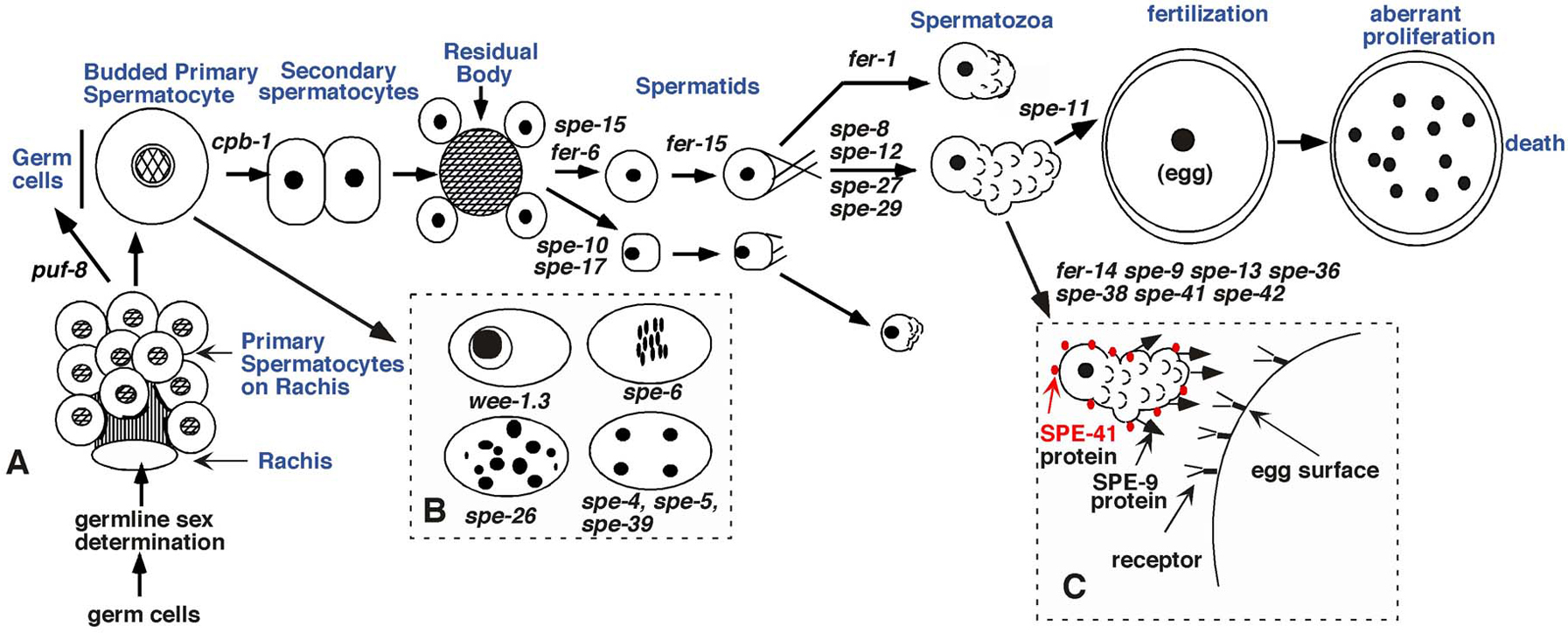
C. elegans spermatogenesis mutants (spe or fer) have been identified because they compromise hermaphrodite self-fertility. In wild-type, virtually every sperm fertilizes an oocyte in young hermaphrodites, and the resulting shelled, oval eggs are laid on the agar growth plate (Ward and Carrel, 1979). In contrast, spe/fer mutants lay unfertilized oocytes, which are round, brown cells that lack an eggshell. The differences between laid oocytes and eggs are easily discerned under the dissecting microscope. Mutant, self-sterile hermaphrodites that have a normal appearing germ line can be readily identified following mutagenesis. While some of these mutants never initiate spermatogenesis (see Sex determination in the germ line), most produce defective sperm. While mutants that fail to initiate spermatogenesis lay a few oocytes, mutants that make defective sperm lay large numbers of unfertilized oocytes (McCarter et al., 1999). Self-sterile hermaphrodites can be mated to wild-type males, allowing the recovery of a spe or fer mutation among the outcross progeny. This straightforward screen has allowed identification of > 44 genes that affect spermatogenesis (reviewed by L'Hernault, 1997; L’Hernault and Singson, 2000; S. L’Hernault, unpublished; S. Ward, unpublished) and several additional genes were identified in other studies (Amiri et al., 2001; Luitjens et al., 2000; Subramaniam and Seydoux, 2003); representative mutants are shown in Figure 5.
|
Figure 5. (A) Stages of wild type spermatogenesis are shown diagramatically as an ordered pathway of morphogenesis with the stages labeled in blue. Some of the > 44 known genes are placed on the pathway where ultrastructural or light microscopic defects are evident. The ife-1 gene is not shown, but it slows down the rate of spermatogenesis and also affects spermatozoa. The next to last step of the pathway represents mutant spermatozoa that are cytologically normal but cannot enter oocytes. The spe-11 mutant is a paternal effect lethal mutant that makes spermatozoa that enter the egg and the resulting defective embryo dies. (B) The most common terminal stages of mutants that arrest morphogenesis without forming spermatids. (C) The protein distribution of SPE-9 (arrows), the hypothesized SPE-9 receptor (“Y” on egg surface) and the SPE-41/TRP-3 protein (red dots). Please note that individual drawings are not to scale; for instance, spermatozoa are 5–6 μm in diameter while the eggs shown are ~45 μm (modified from L'Hernault et al., 1988).
Translational control is widely employed in differentiating cellular systems (reviewed by Gebauer and Hentze, 2004), and two genes have been identified that act as spermatogenesis-specific regulators of translation.
The cpb-1 gene encodes a cytoplasmic polyadenylation element binding protein. Although this gene is expressed in both the testes and the ovary, RNAi experiments reveal that it is required for spermatogenesis but not oogenesis. cpb-1(RNAi) hermaphrodites lay unfertilized oocytes, like spe/fer mutants. Examination of the spermatheca reveals that cpb-1(RNAi) hermaphrodites accumulate spermatocytes and many of these cells fail to complete meiosis. These results suggest that cpb-1 regulates the translation of one or more mRNAs that are critical for completion of meiosis and perhaps other processes during spermatogenesis (Luitjens et al., 2000).
The ife-1 gene encodes an eIF4E mRNA cap binding protein that is associated with P granules during germline proliferation. RNAi of the ife-1 gene results in a temperature sensitive delay in the completion of spermatogenesis, but the majority of the resulting sperm are not competent to engage in fertilization. About 80% of ife-1(RNAi) treated hermaphrodites are self-sterile and the remaining 20% have reduced broods. These data suggest that IFE-1 regulates translation of one or more RNA that are critical for spermatogenesis and sperm function (Amiri et al., 2001).
Like other animals, C. elegans meiosis occurs during both oogenesis and spermatogenesis, and many meiotic processes are similar in the two germ lines. However, two genes that are involved in meiosis can show specific defects during spermatogenesis.
Six dominant wee-1.3(gf) Spe mutants have been discovered and they have no evident defects during oogenesis or in any proliferating somatic tissue. This observation is surprising because Wee1p type kinase is widely expressed (A. Golden, personal communication; Lamitina and L'Hernault, 2002) and probably negatively regulates the C. elegans cdc2p ortholog ncc-1 (Boxem et al., 1999); cdc2p (or its orthologs) is a master regulator of both mitotic and meiotic divisions in eukaryotes (Lundgren et al., 1991). The wee-1.3 null phenotype is embryonic/larval lethality, which indicates it plays an essential role in addition to its role during spermatogenesis (Lamitina and L'Hernault, 2002). These independently obtained dominant wee-1.3 mutations are all located in a four amino acid region near the C-terminus and three contain identical mutations. Dominant wee-1.3 mutant spermatocytes enter pachytene of meiosis I, but the nuclear envelope remains intact and the chromatin acquires an ultrastructural appearance that is similar (although ~4X larger) to that seen in wild-type (haploid) spermatids. wee-1.3 dominant mutants do not initiate cytokinesis and they arrest with an undivided nucleus on one side of the primary spermatocyte (Figure 5B) and vacuolated FB-MOs on the opposite side. These data indicate that a sperm-specific pathway negatively regulates WEE-1.3 during the G2/M transition of male meiosis I, and this regulation does not occur in these dominant mutants (Lamitina and L'Hernault, 2002).
puf-8 encodes a pumilio-like RNA binding protein that controls RNA stability and translation. The PUF-8 protein is required redundantly with other PUF proteins to maintain viable germ cells during development (Subramaniam and Seydoux, 1999), but it also plays a partially penetrant, non redundant role in the testes. Primary spermatocytes lacking PUF-8 complete prophase of meiosis I, but then exit meiosis, re-enter mitosis and de-differentiate into tumorous germ cells. This observation indicates that PUF-8 is required for primary spermatocytes to complete meiosis and remain in the spermatogenesis pathway (Subramaniam and Seydoux, 2003).
Several mutants affect FB-MO morphogenesis or function.
The spe-39 gene encodes a novel hydrophilic, cytoplasmic protein that is not specifically associated with FB-MOs in spermatocytes and spermatids. spe-39 mutants arrest as aberrant spermatocytes (Figure 5B) that lack MOs but contain many small vesicles with internal membranes, a distinctive feature of wild-type MOs (Figure 1). A membrane envelope does not surround spe-39 FBs, indicating that some aspects of the FB can form in the absence of MOs. In addition to its role during spermatogenesis, SPE-39 is found in many, if not all, C. elegans cells, and spe-39 RNAi causes embryonic lethality. The spe-39 gene has orthologs in many metazoa, but no homolog was identified in the yeast genome. This observation suggests that the SPE-39 role during FB-MO morphogenesis is a conserved process that requires SPE-39 orthologs in other metazoan cell types (Zhu and L'Hernault, 2003).
The spe-6 gene encodes a casein I type serine threonine kinase (Muhlrad and Ward, 2002). In spe-6 null mutants, the MSP fails to form FBs (Varkey et al., 1993). Consequently, FB-MOs never form, so spe-6 mutants do not form spermatids and arrest as aberrant spermatocytes (Figure 5B). This observation suggests that a regulatory phosphorylation controls FB morphogenesis, but the substrates for the SPE-6 kinase are not currently defined. Non-null mutants reveal that this gene is also involved in regulating spermiogenesis (see Sex-specific aspects of spermiogenesis, below).
The spe-4 gene encodes a sperm-specific presenilin (L'Hernault and Arduengo, 1992); presenilins are intramembranous aspartyl proteases that process other membrane proteins, including the Alzheimer precursor protein in human brain (Xia and Wolfe, 2003). The SPE-4 protein resides in FB-MOs and spe-4 mutants, like spe-39 mutants, develop FBs that are not associated with MOs. spe-4 mutants accumulate aberrant spermatocytes filled with distended MOs that are not associated with FBs (Arduengo et al., 1998).
The spe-5 gene encodes a vacuolar ATPase B subunit (P. Hartley and S.W. L’Hernault, unpublished) that localizes to the FB-MO (E. J. Gleason and S. W. L’Hernault, unpublished). spe-5 mutants form FB-MOs that are greatly distended and vacuolated. Usually, spe-5 null mutants arrest as aberrant spermatocytes that contain four haploid nuclei (Figure 5B), but occasionally a few spe-5 self-progeny are observed so some functional spermatozoa must form (Machaca and L'Hernault, 1997). Because vacuolar ATPases acidify compartments (Nishi and Forgac, 2002), the spe-5 defect may reflect a lack of FB-MO acidification.
The spe-17 gene encodes a small soluble protein with no obvious conserved domains or homologs outside nematodes (L'Hernault et al., 1993). spe-17 mutants form FB-MOs with membrane-attached ribosomes. The consequence is that spe-17 mutants retain ribosomes in the spermatid. The FB can still disassemble and release MSP, but many MOs do not fuse with the cell surface during spermiogenesis (Shakes and Ward, 1989). Motile spermatozoa can still form in spe-17 mutants, and some are competent to engage in fertilization.
spe-10 mutants initiate FB-MO morphogenesis normally, but the membrane surrounding the FB prematurely retracts so that FBs are left in the residual body during spermatid budding. Budded spe-10 spermatids are deficient in MSP and resulting spe-10 spermatozoa are immotile and have vacuolated MOs that do not fuse with the plasma membrane (Shakes and Ward, 1989). The spe-10 gene encodes a four pass integral membrane protein that contains a DHHC domain predicted to be a palmitoyl transferase that is localizes within FB-MOs (Gleason et al. 2006). The presence of this motif implicates SPE-10 function in the posttranslational palmitoylation of target proteins required for proper FB-MO function. In addition to FB-MO defects, both spe-17 and spe-10 mutant males make spermatids that are only ~66% the wild-type size and have nuclei that are eccentrically placed (Shakes and Ward, 1989).
fer-6 mutants makes spermatids in which the FBs detach from the MO but fail to disassemble and release MSP. Many MOs fail to fuse with the plasma membrane in fer-6 mutants, which form abnormal spermatids (Ward et al., 1981).
fer-15 mutant spermatids fail to respond to in vitro activators and do not become spermatozoa (Figure 5; S. L’Hernault and S. Ward, unpublished observations).
fer-1 mutants have no apparent defects in the early stages of FB-MO morphogenesis and have normal sized spermatids in which FBs disassemble normally. However, the MOs fail to fuse with the plasma membrane, and fer-1 mutants extend short pseudopods that do not allow normal motility (Roberts and Ward, 1982; Ward et al., 1981). The fer-1 gene encodes a one pass transmembrane protein in the dysferlin family with homology to at least seven human genes (Achanzar and Ward, 1997), including one implicated in limb girdle muscular dystrophy (Bashir et al., 1998). Human dysferlins are probably involved in muscle membrane repair/remodeling (Bansal and Campbell, 2004), which is consistent with the fer-1 membrane fusion defect (Ward et al., 1981).
There are two spe genes that encode known cytoskeletal proteins.
The spe-26 gene encodes an actin binding protein, and mutants usually form aberrant spermatocytes that do not become spermatids. Occasionally, spe-26 mutants make spermatids that become spermatozoa, but these spermatozoa do not participate in fertilization. A spe-26 spermatocyte can complete meiosis and form four haploid nuclei within a single, aberrant cell, but these cells sometimes contain as many as 12 DAPI positive DNA regions (Varkey et al., 1995).
The spe-15 gene encodes a myosin VI that plays a sorting role as spermatids bud from the residual body. spe-15 mutants partially fail in their polarized delivery of mitochondria and FB-MOs to spermatids and actin filaments and microtubules to the residual body. As a consequence, all of these cellular components are distributed to both spermatids and residual bodies during budding. When spe-15 spermatids are protease activated in vitro, resulting spermatozoa usually lack normal pseudopods (Kelleher et al., 2000).
Sperm derived from hermaphrodites and males have two known differences. Male-derived sperm are substantially bigger than hermaphrodite-derived sperm (Figure 3; LaMunyon and Ward, 1995; LaMunyon and Ward, 1998), and the regulatory machinery they utilize for spermiogenesis is sex-specific. Mutant screens for spermatogenesis defects have uncovered four genes (the spe-8 pathway) with hermaphrodite-specific spermiogenesis defects (L'Hernault et al., 1988; Minniti et al., 1996; Nance et al., 1999; Nance et al., 2000; Shakes and Ward, 1989).
spe-8 encodes a predicted SH2 domain containing non receptor tyrosine kinase (P.J. Muhlrad and S. Ward, unpublished).
spe-12 encodes a single pass transmembrane protein. This novel protein is predicted to be on the sperm surface (Nance et al., 1999).
spe-27 encodes a ~15 kD protein that is novel and hydrophilic (Minniti et al., 1996).
spe-29 encodes a single pass transmembrane protein that is only ~7.5 kD. This novel protein is predicted to be on the sperm surface (Nance et al., 2000).
Hermaphrodites homozygous for a mutation in any spe-8 pathway gene accumulate spermatids that fail to activate into spermatozoa (Figure 5). Unlike most (> 40) spermatogenesis genes, males homozygous for a mutation in any one of these four genes produce spermatozoa that are competent for fertilization. The arrested spermatids present in hermaphrodites that are mutant in the spe-8 pathway can be activated to become spermatozoa if they are exposed to male-derived seminal fluid. However, neither male nor hermaphrodite derived spermatids from either sex are entirely normal; after protease treatment, mutant spermatids have an unusual spiky cytology, whereas wild-type spermatids become cytologically normal spermatozoa (Minniti et al., 1996; Nance et al., 1999; Nance et al., 2000; Shakes and Ward, 1989).
Eighteen non-null spe-6 mutants allow partial bypass of any spe-8 pathway mutant (spe-6 null mutants have defects in FB-MO morphogenesis, see above). These non-null spe-6 mutations are scattered across the polypeptide sequence and some affect non-essential residues in the kinase catalytic region. It is thought that SPE-6 negatively regulates spermiogenesis until an activation signal is sent through the spe-8 pathway. Partial loss of SPE-6 function partly alleviates the need for spe-8 pathway-mediated relief of SPE-6 negative regulation in hermaphrodites (Muhlrad and Ward, 2002). These spe-6 suppressor mutants have also been analyzed in a background that was not mutant for other genes in the spe-8 pathway. In males, spe-6 suppressor mutations show precocious activation of spermatids into spermatozoa. However, there is a large reduction in spermatid numbers due to necrosis, and mutant males rarely sire progeny after copulation. In contrast, hermaphrodite-derived spermatids show only minimal signs of necrosis in spe-6 suppressor mutants (Muhlrad and Ward, 2002).
These data indicate that there are two spermiogenesis pathways, one active in males and one active in hermaphrodites (Nance et al., 2000; Shakes, 1988). The pathway in hermaphrodites has two kinases (SPE-6 and SPE-8), one soluble protein (SPE-27) and two transmembrane proteins (SPE-12 and SPE-29), suggesting it functions in signal transduction. The physiological signal that initiates spermiogenesis in hermaphrodites is not presently known. None of the genes that participate in the spermiogenesis pathway for male-derived spermatids are presently known.
Seven mutants (fer-14, spe-9, spe-13, spe-36, spe-38, spe-41/trp-3 and spe-42) appear to complete spermatogenesis normally but are defective in fertilization (Figure 5; also see Control of oocyte meiotic maturation and fertilization), and five of these genes have been cloned.
The spe-9 gene encodes a one-pass integral membrane protein with 10 EGF-like motifs, which usually have extracellular functions such as adhesive and ligand-receptor interactions (Singson et al., 1999). Transgenes bearing the spe-9 coding sequence in which deletion or point mutations were induced revealed that certain EGF repeats in the predicted SPE-9 protein are more important than others for SPE-9 function during fertilization (Putiri et al., 2004).
The spe-41/trp-3 gene encodes a TRP calcium channel protein that has a role in store-and, perhaps, receptor- operated calcium entry during fertilization (Xu and Sternberg, 2003).
The spe-38 gene encodes a novel tetraspan integral membrane protein (Chatterjee et al., 2005). Structurally similar proteins participate in membrane adhesion/fusion during mammalian fertilization, yeast mating and metazoan tight junction formation (Hemler, 2003), but the precise role of SPE-38 remains to be determined.
The fer-14 gene encodes a novel one pass transmembrane protein that is nematode specific. (T. Kroft, E. J. Gleason and S. W. L’Hernault, unpublished).
The spe-42 gene encodes a seven pass transmembrane protein, and two SPE-42 homologs are evident in all multicellular animals with a complete genome sequence, including C. elegans. In humans, one of these homologs has a splice form expressed in testes, but the precise function of SPE-42 or any of its homologs is not yet known (Kroft et al., 2005).
Spermatozoa derived from any of the seven mutants in this class have no detectable motility defects, and they make contact with oocytes in the spermatheca. Male-derived spermatozoa from five of these mutants (fer-14, spe-9, spe-13, spe-41/trp-3 and spe-42) render recipient, formerly self-fertile, hermaphrodites sterile after mating; such hermaphrodites cease laying embryos and begin to lay unfertilized oocytes (Singson et al., 1999; Xu and Sternberg, 2003). This sterility reflects the fact that these mutant, male-derived spermatozoa can out-compete and functionally displace endogenous, fertilization-competent hermaphrodite spermatozoa in the spermatheca (as in Figure 5C). The consequence of this displacement is that oocytes only contact fertilization defective spermatozoa during the narrow temporal window when they are competent for fertilization.
SPE-41 (Xu and Sternberg, 2003), SPE-9 (Zannoni et al., 2003) and SPE-38 (Chatterjee et al., 2005) are all plasma membrane proteins in spermatozoa. SPE-9 is also found in the plasma membrane of spermatids, but SPE-41 and SPE-38 are found in the MOs and become plasma membrane proteins when the MOs fuse during spermiogenesis. This observation places the SPE-41 TRP channel function on the cell surface where it could participate in calcium fluxes related to gamete fusion during fertilization. While SPE-41 is distributed on the plasma membrane of both the cell body and the pseudopod, SPE-9 and SPE-38 localize to the pseudopodial plasma membrane of spermatozoa (Figure 5C). These data indicate that there are at least two ways for plasma membrane proteins required for fertilization to reach the spermatozoan cell surface. Additional proteins important for oocyte recognition, adhesion and/or fusion are likely to be encoded by spe-13 and spe-36 and other sperm-expressed genes that remain to be discovered.
One mutant (spe-11) forms spermatozoa that are competent to fertilize oocytes, but embryogenesis never begins properly and the defective embryo always dies (Browning and Strome, 1996; Hill et al., 1989; L'Hernault et al., 1988). After a wild type oocyte is fertilized by a spe-11 spermatozoon, maternal meiosis is not usually completed properly and a defective eggshell forms. While mitosis is observed in oocytes fertilized by a spe-11 sperm, the spindle is not positioned correctly and cytokinesis does not usually occur. The typical end result is that multiple nuclei share a common cytoplasm in these defective embryos (Figure 5; Hill et al., 1989). Expression of the spe-11 gene normally occurs only during spermatogenesis, and the SPE-11 protein has no clear homologs outside nematodes (Browning and Strome, 1996). Ultrastructural examination of spe-11 mutant spermatids reveals that they have defects in perinuclear material surrounding the nucleus (Hill et al., 1989), which is where the SPE-11 protein localizes in wild type spermatids (Browning and Strome, 1996). This sperm-derived material is apparently required for some as yet unknown aspect of early embryogenesis. However, it does not have to be delivered via the sperm because transgenic expression in the oocyte is sufficient to allow embryos that have been fertilized with a spe-11 spermatozoon to survive (Browning and Strome, 1996). Other work has shown that the C. elegans sperm-derived nucleus (Sadler and Shakes, 2000) and centriole (O'Connell, 2002; O'Connell et al., 2001) are both required for embryogenesis. However, SPE-11 is the only known C. elegans protein that is both required for embryogenesis and normally has a sperm-specific expression pattern.
Progress in the study of C. elegans spermatogenesis has accelerated in recent years as the molecular identities of many of the involved genes have been discovered. The earliest stages of spermatogenesis employ genes (cpb-1, ife-1, puf-8, spe-39 and wee-1.3) that are not exclusively expressed during spermatogenesis. If genes exist that regulate translational control or meiosis, but commence expression after spermatogenesis has been initiated, they remain to be discovered. A partial picture of the function of the FB-MO has emerged, but much work remains to be done. The earliest stages of FB-MO morphogenesis are presumably controlled through the vesicular trafficking pathway. These components are likely to be expressed outside the testes (as is spe-39) and probably will require different types of mutant screens and/or RNAi studies to be identified. Although the genetic control of hermaphrodite spermiogenesis is becoming clearer, none of the genes that participate in the predicted male spermiogenesis pathway have been identified. Mutants with specific defects in male spermiogenesis are likely obtainable, but existing screening methods that rely on recovering self-sterile hermaphrodites will not allow identification of such mutants; new screens will need to be designed for this purpose. There has been much recent progress in identifying and analyzing the sperm-expressed genes that participate in fertilization. All five of the cloned genes that are important for fertilization encode transmembrane proteins. How these transmembrane proteins organize to interact with the egg during fertilization remains to be determined. The spe-11 mutant reveals that sperm play a crucial role in the delivery of components to the oocyte that allow it to progress through embryogenesis normally. It will be interesting to learn if there are other proteins required for embryogenesis that must be delivered by the sperm during fertilization.
I thank Sam Ward, Andrew Singson and members of the L’Herrnault lab for valuable discussion. Work in my laboratory has been supported by grants from the NIH (GM040697) and NSF (IBN-0131532).
Achanzar, W.E., and Ward, S. (1997). A nematode gene required for sperm vesicle fusion. J. Cell Sci. 110, 1073–1081. Abstract
Amiri, A., Keiper, B.D., Kawasaki, I., Fan, Y., Kohara, Y., Rhoads, R.E., and Strome, S. (2001). An isoform of eIF4E is a component of germ granules and is required for spermatogenesis in C. elegans. Development 128, 3899–3912. Abstract
Arduengo, P.M., Appleberry, O.K., Chuang, P., and L'Hernault, S.W. (1998). The presenilin protein family member SPE-4 localizes to an ER/Golgi derived organelle and is required for proper cytoplasmic partitioning during Caenorhabditis elegans spermatogenesis. J. Cell. Sci. 111, 3645–3654. Abstract
Bansal, D., and Campbell, K.P. (2004). Dysferlin and the plasma membrane repair in muscular dystrophy. Trends Cell Biol. 14, 206–213. Abstract Article
Bashir, R., Britton, S., Strachan, T., Keers, S., Vafiadaki, E., Lako, M., Richard, I., Marchand, S., Bourg, N., Argov, Z., et al. (1998). A gene related to Caenorhabditis elegans spermatogenesis factor fer-1 is mutated in limb-girdle muscular dystrophy type 2B. Nat. Genet. 20, 37–42. Abstract Article
Boxem, M., Srinivasan, D.G., and van den Heuvel, S. (1999). The Caenorhabditis elegans gene ncc-1 encodes a cdc2-related kinase required for M phase in meiotic and mitotic cell divisions, but not for S phase. Development 126, 2227–2239 Abstract
Browning, H., and Strome, S. (1996). A sperm-supplied factor required for embryogenesis in C. elegans. Development 122, 391–404. Abstract
Chatterjee, I., Richmond, A., Putiri, E., Shakes, D.C., and Singson, A. (2005). The Caenorhabditis elegans spe-38 gene encodes a novel four-pass integral membrane protein required for sperm function at fertilization. Development 132, 2795–2808. Abstract Article
Gebauer, F., and Hentze, M.W. (2004). Molecular mechanisms of translational control. Nat. Rev. Mol. Cell Biol. 5, 827–835. Abstract Article
Gleason, E.J., Lindsey, W.C., Kroft, T.L., Singson, A.W., and L'hernault, S.W. (2006). spe-10 encodes a DHHC-CRD zinc-finger membrane protein required for endoplasmic reticulum/Golgi membrane morphogenesis during Caenorhabditis elegans spermatogenesis. Genetics 172, 145–158. Abstract Article
Hemler, M.E. (2003). Tetraspanin proteins mediate cellular penetration, invasion, and fusion events and define a novel type of membrane microdomain. Annu. Rev. Cell Dev. Biol. 19, 397–422. Abstract Article
Hill, D.P., Shakes, D.C., Ward, S., and Strome, S. (1989). A sperm-supplied product essential for initiation of normal embryogenesis in Caenorhabditis elegans is encoded by the paternal-effect embryonic-lethal gene, spe-11. Dev. Biol. 136, 154–166. Abstract Article
Kelleher, J.F., Mandell, M.A., Moulder, G., Hill, K.L., L'Hernault, S.W., Barstead, R., and Titus, M.A. (2000). Myosin VI is required for asymmetric segregation of cellular components during C. elegans spermatogenesis. Curr. Biol. 10, 1489–1496. Abstract Article
Kimble, J., and Ward, S. (1988). Germ-Line Development and Fertilization. In The Nematode Caenorhabditis elegans, W.B. Wood, ed. (Cold Spring Harbor, Cold Spring Harbor Laboratory Press), pp. 191–213.
Klass, M.R., and Hirsh, D. (1981). Sperm isolation and biochemical analysis of the major sperm protein from Caenorhabditis elegans. Dev. Biol. 84, 299–312. Article
Kroft, T.L., Gleason, E.J., and L'Hernault, S.W. (2005). The spe-42 gene is required for sperm-egg interactions during C. elegans fertilization and encodes a sperm-specific transmembrane protein. Dev. Biol. 286, 169–181. Abstract Article
Lamitina, S.T., and L'Hernault, S.W. (2002). Dominant mutations in the Caenorhabditis elegans Myt1 ortholog wee-1.3 reveal a novel domain that controls M-phase entry during spermatogenesis. Development 129, 5009–5018. Abstract
LaMunyon, C.W., and Ward, S. (1994). Assessing the viability of mutant and manipulated sperm by artificial insemination of Caenorhabditis elegans. Genetics 138, 689–692. Abstract
LaMunyon, C.W., and Ward, S. (1995). Sperm precedence in a hermaphroditic nematode (Caenorhabditis elegans) is due to competitive superiority of male sperm. Experientia 51, 817–823. Abstract Article
LaMunyon, C.W., and Ward, S. (1998). Larger sperm outcompete smaller sperm in the nematode Caenorhabditis elegans. Proc. R. Soc. Lond. B Biol. Sci. 265, 1997–2002. Abstract Article
L'Hernault, S.W. (1997). Male germ line. In C. elegans II, D. Riddle, R. Blumenthal, B.J. Meyer, and J. Priess, eds. (Cold Spring Harbor, NY: Cold Spring Harbor Press), pp. 271–294.
L'Hernault, S.W., and Arduengo, P.M. (1992). Mutation of a putative sperm membrane protein in Caenorhabditis elegans prevents sperm differentiation but not its associated meiotic divisions. J. Cell Biol. 119, 55–68. Abstract Article
L'Hernault, S.W., Benian, G.M., and Emmons, R.B. (1993). Genetic and molecular characterization of the Caenorhabditis elegans spermatogenesis-defective gene spe-17. Genetics 134, 769–780. Abstract
L'Hernault, S.W., Shakes, D.C., and Ward, S. (1988). Developmental genetics of chromosome I spermatogenesis-defective mutants in the nematode Caenorhabditis elegans. Genetics 120, 435–452. Abstract
Luitjens, C., Gallegos, M., Kraemer, B., Kimble, J., and Wickens, M. (2000). CPEB proteins control two key steps in spermatogenesis in C. elegans. Genes Dev. 14, 2596–2609. Abstract Article
Lundgren, K., Walworth, N., Booher, R., Dembski, M., Kirschner, M., and Beach, D. (1991). mik1 and wee1 cooperate in the inhibitory tyrosine phosphorylation of cdc2. Cell 64, 1111–1122. Abstract Article
L'Hernault, S.W., and Singson, A.W. (2000). Developmental Genetics of Spermatogenesis in the Nematode Caenorhabditis elegans. In The Testes: From Stem Cell to Sperm Function, Serono Symposium USA, E. Goldberg, ed. (New York, Springer-Verlag), pp. 109–119.
Machaca, K., DeFelice, L.J., and L'Hernault, S.W. (1996). A novel chloride channel localizes to Caenorhabditis elegans spermatids and chloride channel blockers induce spermatid differentiation. Dev. Biol. 176, 1–16. Abstract Article
Machaca, K., and L'Hernault, S.W. (1997). The Caenorhabditis elegans spe-5 gene is required for morphogenesis of a sperm-specific organelle and is associated with an inherent cold-sensitive phenotype. Genetics 146, 567–581. Abstract
McCarter, J., Bartlett, B., Dang, T., and Schedl, T. (1999). On the control of oocyte meiotic maturation and ovulation in Caenorhabditis elegans. Dev. Biol. 205, 111–128. Abstract Article
Minniti, A.N., Sadler, C., and Ward, S. (1996). Genetic and molecular analysis of spe-27, a gene required for spermiogenesis in Caenorhabditis elegans hermaphrodites. Genetics 143, 213–223. Abstract
Muhlrad, P.J., and Ward, S. (2002). Spermiogenesis initiation in Caenorhabditis elegans involves a casein kinase 1 encoded by the spe-6 gene. Genetics 161, 143–155. Abstract
Nance, J., Davis, E.B., and Ward, S. (2000). spe-29 encodes a small predicted membrane protein required for the initiation of sperm activation in Caenorhabditis elegans. Genetics 156, 1623–1633. Abstract
Nance, J., Minniti, A.N., Sadler, C., and Ward, S. (1999). spe-12 encodes a sperm cell surface protein that promotes spermiogenesis in Caenorhabditis elegans. Genetics 152, 209–220. Abstract
Nelson, G.A., Roberts, T.M., and Ward, S. (1982). Caenorhabditis elegans spermatozoan locomotion: amoeboid movement with almost no actin. J. Cell Biol. 92, 121–131. Abstract Article
Nelson, G.A., and Ward, S. (1980). Vesicle fusion, pseudopod extension and amoeboid motility are induced in nematode spermatids by the ionophore monensin. Cell 19, 457–464. Abstract Article
Nishi, T., and Forgac, M. (2002). The vacuolar (H+)-ATPases--nature's most versatile proton pumps. Nat. Rev. Mol. Cell Biol. 3, 94–103. Abstract Article
O'Connell, K.F. (2002). The ZYG-1 kinase, a mitotic and meiotic regulator of centriole replication. Oncogene 21, 6201–6208. Abstract Article
O'Connell, K.F., Caron, C., Kopish, K.R., Hurd, D.D., Kemphues, K.J., Li, Y., and White, J.G. (2001). The C. elegans zyg-1 gene encodes a regulator of centrosome duplication with distinct maternal and paternal roles in the embryo. Cell 105, 547–558. Abstract Article
Putiri, E., Zannoni, S., Kadandale, P., and Singson, A. (2004). Functional domains and temperature-sensitive mutations in SPE-9, an EGF repeat-containing protein required for fertility in Caenorhabditis elegans. Dev. Biol. 272, 448–459. Abstract Article
Roberts, T.M., Pavalko, F.M., and Ward, S. (1986). Membrane and cytoplasmic proteins are transported in the same organelle complex during nematode spermatogenesis. J. Cell Biol. 102, 1787–1796. Abstract Article
Roberts, T.M., and Ward, S. (1982). Centripetal flow of pseudopodial surface components could propel the amoeboid movement of Caenorhabditis elegans spermatozoa. J. Cell Biol. 92, 132–138. Abstract Article
Sadler, P.L., and Shakes, D.C. (2000). Anucleate Caenorhabditis elegans sperm can crawl, fertilize oocytes and direct anterior-posterior polarization of the 1-cell embryo. Development 127, 355–366. Abstract
Shakes, D.C. (1988) A genetic and pharmacological analysis of spermatogenesis in the nematode, Caenorhabditis elegans.Ph.D. Thesis, Johns Hopkins University, Baltimore.
Shakes, D.C., and Ward, S. (1989). Initiation of spermiogenesis in C. elegans: a pharmacological and genetic analysis. Dev. Biol. 134, 189–200. Abstract Article
Shakes, D.C., and Ward, S. (1989). Mutations that disrupt the morphogenesis and localization of a sperm-specific organelle in Caenorhabditis elegans. Dev. Biol. 134, 307–316. Article
Singson, A., Hill, K.L., and L'Hernault, S.W. (1999). Sperm competition in the absence of fertilization in Caenorhabditis elegans. Genetics 152, 201–208. Abstract
Subramaniam, K., and Seydoux, G. (1999). nos-1 and nos-2, two genes related to Drosophila nanos, regulate primordial germ cell development and survival in Caenorhabditis elegans. Development 126, 4861–4871. Abstract
Subramaniam, K., and Seydoux, G. (2003). Dedifferentiation of primary spermatocytes into germ cell tumors in C. elegans lacking the pumilio-like protein PUF-8. Curr. Biol. 13, 134–139. Abstract Article
Varkey, J.P., Jansma, P.L., Minniti, A.N., and Ward, S. (1993). The Caenorhabditis elegans spe-6 gene is required for major sperm protein assembly and shows second site non-complementation with an unlinked deficiency. Genetics 133, 79–86. Abstract
Varkey, J.P., Muhlrad, P.J., Minniti, A.N., Do, B., and Ward, S. (1995). The Caenorhabditis elegans spe-26 gene is necessary to form spermatids and encodes a protein similar to the actin-associated proteins kelch and scruin. Genes Dev. 9, 1074–1086. Abstract
Ward, S. (1986). The asymmetric localization of gene products during the development of Caenorhabditis elegans spermatozoa. In Gametogenesis and the Early Embryo, J. Gall, ed. (New York: A.R. Liss), pp. 55–75.
Ward, S., Argon, Y., and Nelson, G.A. (1981). Sperm morphogenesis in wild-type and fertilization-defective mutants of Caenorhabditis elegans. J. Cell Biol. 91, 26–44. Abstract Article
Ward, S., and Carrel, J.S. (1979). Fertilization and sperm competition in the nematode Caenorhabditis elegans. Dev. Biol. 73, 304–321. Abstract
Ward, S., Hogan, E., and Nelson, G.A. (1983). The initiation of spermiogenesis in the nematode Caenorhabditis elegans. Dev. Biol. 98, 70–79. Abstract Article
Ward, S., and Klass, M. (1982). The location of the major protein in Caenorhabditis elegans sperm and spermatocytes. Dev. Biol. 92, 203–208. Abstract Article
Wolf, N., Hirsh, D., and McIntosh, J.R. (1978). Spermatogenesis in males of the free-living nematode, Caenorhabditis elegans. J. Ultrastruct. Res. 63, 155–169. Abstract Article
Xia, W., and Wolfe, M.S. (2003). Intramembrane proteolysis by presenilin and presenilin-like proteases. J. Cell Sci. 116, 2839–2844. Abstract Article
Xu, X.Z., and Sternberg, P.W. (2003). A C. elegans sperm TRP protein required for sperm-egg interactions during fertilization. Cell 114, 285–297. Abstract Article
*Edited by Judith Kimble and Susan Strome. Last revised March 14, 2005. Published February 20, 2006. This chapter should be cited as: L'Hernault, S.W. Spermatogenesis (February 20, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.85.1, http://www.wormbook.org.
Copyright: © 2006 S.W. L'Hernault. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: bioslh@biology.emory.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.