Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
The easiest and most efficient way to recover Caenorhabditis and some other nematodes from soil or any other substrate is to place the sample onto a standard C. elegans culture plate (see section 2). Alternative methods used by nematologists to recover soil nematodes (see sections 3 and 4) are in our hands more difficult to implement and only yield a fraction of the individuals in the sample. More detailed information on different sampling methods can be found in Hooper (1986) or De Ley and Mundo-Ocampo (2004). The most difficult part is then to recognize your species of interest out of the zoo of species that comes with a typical soil or compost sample (see section 5). We summarize the organization and timing of an isolation experiment (see section 6). Finally, culture (see section 7) and freezing (see section 8) conditions are briefly reviewed. Bear in mind that this chapter is strongly biased towards the isolation of Caenorhabditis elegans and close relatives.
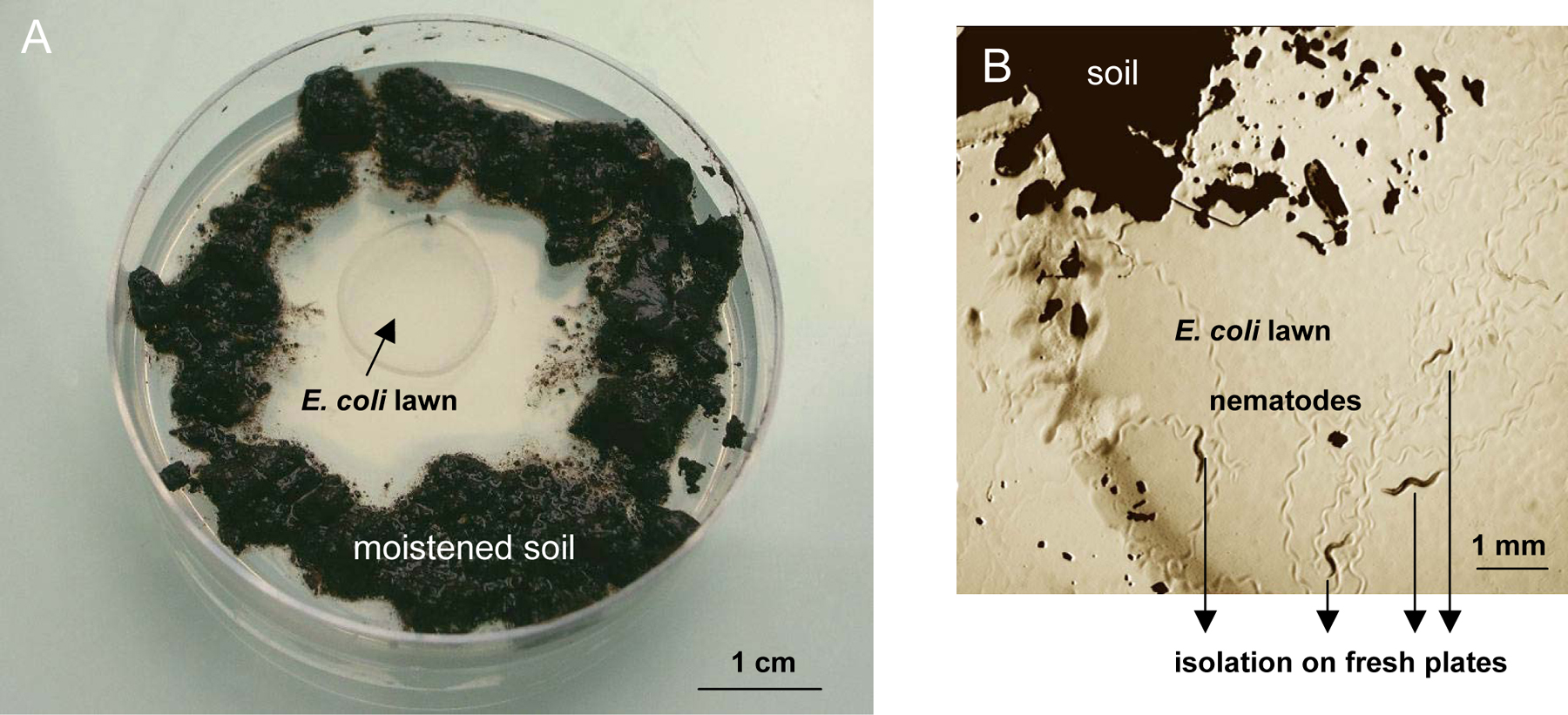
The easiest way to recover nematodes from soil is simply to place the substrate around the bacterial lawn of a standard C. elegans culture dish (see Maintenance of C. elegans). Bacteria-feeding nematodes such as C. elegans are attracted and crawl out of the sample towards the bacterial lawn (Figure 1). For fast-moving species like C. elegans, this occurs within minutes to hours.
|
Figure 1. Nematode isolation on an agar plate. A. The substrate (soil, compost, etc.) is placed around the E. coli lawn onto a standard ‘C. elegans’ culture plate. B. Individual nematodes that crawl out can be picked onto new culture plates.
Spread about 1–2 grams of soil around the E. coli OP50 lawn of a standard NGM Petri dish, and moisten with about 1 ml M9 solution (3 g KH2PO4, 6 g Na2HPO4, 5 g NaCl, 1 ml 1 M MgSO4, H2O to 1 liter. Sterilize by autoclaving.) or water.
Pick individual worms that have crawled out to a new plate (in obligate male-female species, worms of both sexes or a mated female are required). Worms are ideally picked within a few hours after the sample has been placed on the plate. Later on, growing bacteria and fungi species that have been introduced with the sample may prevent worms from accessing the OP50 lawn (to avoid such growth, the sample can be moistened with 0.01% thimerosal in M9).
Using this method, all developmental stages can be recovered from soil and compost samples in a quantitative manner (except for the embryonic stages, which can only be retrieved after hatching). The number of individuals in the original sample can be measured in this way (Barrière and Félix, 2005). Some nematode species, including Caenorhabditis elegans, will eventually form dauers that present a characteristic waving behavior: they ‘stand’ on their tail on any substrate with a narrow tip (the substrate or the molds that grow out of it) and move their body, putatively as a way to be carried away by an invertebrate. One can thus a posteriori check that every Caenorhabditis individual has been picked by the absence of waving dauer larvae after 6–10 days.
Many nematode species are associated with invertebrates (molluscs, arthropods, etc.) during the dauer stage (see Ecology of Caenorhabditis species; Kiontke, 1997). They can be recovered by sacrifying the carriers, placing them on NGM plates and waiting for the worms to move to the OP50 lawn or collect them directly from the decaying invertebrate host.
C. elegans has been isolated from compost heaps (decaying leaves or vegetal matter), garden soil and carrier invertebrates (Barrière and Félix, 2005; Cutter, 2006; Haber et al., 2005; Sivasundar and Hey, 2005). Recently, we also found large populations of Caenorhabditis species in fruits decaying under their tree (figs, apples, etc.).
This is the classical method to recover all species of nematodes, particularly those not attracted by E. coli. Many modifications and improvements are possible, and here we show only a single basic design.
Fit a rubber tube onto the stem of a funnel (diameter ca. 50 mm). Close the tube with a clamp (a simple paperclip can be used instead) and fill the rubber tubing with M9 solution up to the neck of the funnel (Figure 2).
Line the funnel cone with a piece of sieve (standard cleaning tissue, muslin, mosquito mesh…), then fill with soil material and fold to enclose the sample. Add M9 solution or water until the sample is submerged.
Active nematodes crawl through the sieve and sediment at the end of the tubing. Recover them by opening the clamp (after a few hours until several days).
|
Figure 2. Baermann funnel. The nematodes swim towards the base of the funnel (this may require several days for some species; rhabditids come out after a few hours). Courtesy of K. Kiontke.
A problem with the Baermann funnel technique, especially when isolating worms from bacteria-rich samples, is the poor oxygenation of the sample that results in nematode mortality.
Terrestrial nematodes have a specific gravity of 1.08 and can be recovered by flotation on a solution with a specific gravity of 1.15 (Hooper, 1986). This method is best used with soil samples, whereas decaying leaves or compost material yield too much detritus.
Several solutions can be used:
Sucrose - inexpensive, but sticky and has a strong osmotic effect that harms the worms (1.71 mol/L for a density of 1.15).
MgSO4 - less sticky than sucrose, but also has a strong osmotic effect (1.38 mol/L).
Ludox - colloidal silica: expensive, but weak osmotic effect; samples can therefore be extracted several times, in contrast to extraction with the other solutions. Ludox comes in two forms: Ludox HS40, toxic, that may be used for fixed samples, and Ludox AS, non-toxic, that is best used to recover live material. A 50% solution in water has a specific gravity of 1.15.
If your sample contains a lot of debris, rinse it first over a 1 mm mesh to remove the bulkiest fragments (this will however remove some of the nematodes).
Soak the sample in water and centrifuge at 1,800 g to obtain a compact pellet, and eliminate the supernatant.
Resuspend the pellet in 20 ml of the chosen solution of density 1.15 and centrifuge at 1,800 g for 1 minute. Add 2 to 5 ml of water or M9 buffer on top and centrifuge again for 5-15 minutes. Nematodes will accumulate at the interface between the two solutions.
Pipet the worms at the interface (some can be seen with the naked eye) using a cut 1 ml pipette tip.
Rinse them in M9, centrifuge, and pipet the pellet onto a standard NGM culture plate.
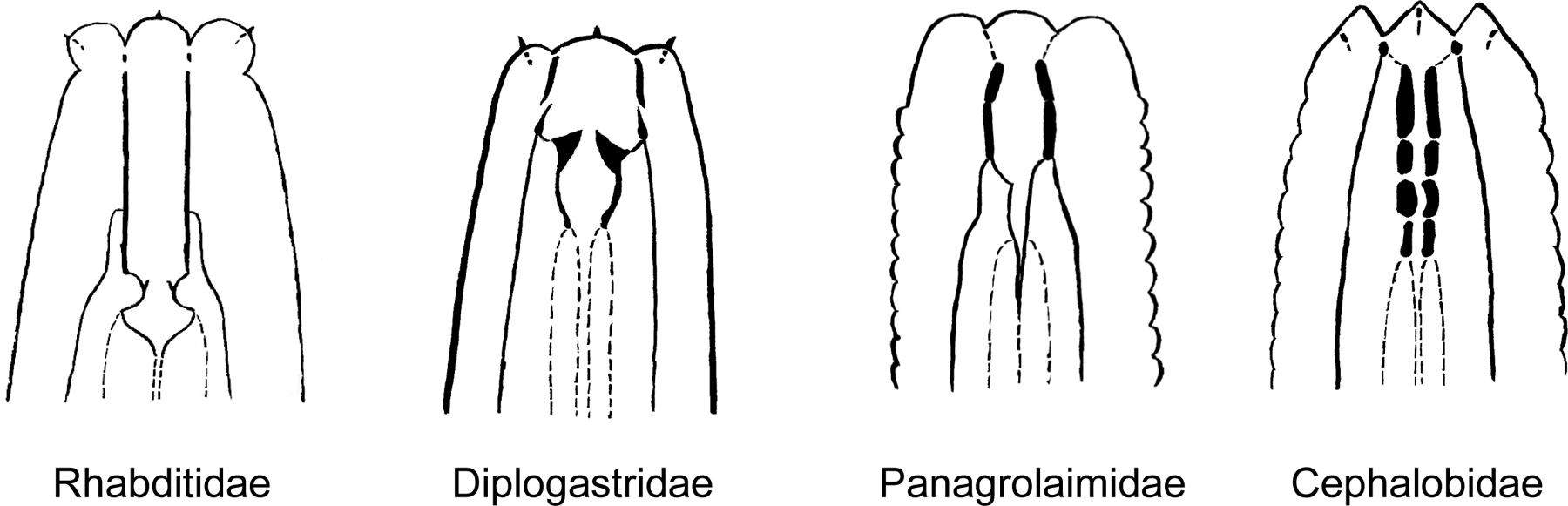
The worms can be identified by morphological criteria either as they come out of the substrate, or on their progeny after they were brought in culture (see section 6). Except for a few plant parasitic nematodes (which are very long nematodes with a stylet in the mouth that helps them invading their hosts) and fungal feeders like aphelenchids, most nematodes from soil or compost samples belong to one of four families: Rhabditidae (including Caenorhabditis, Oscheius, Mesorhabditis, etc.), Diplogastridae (Pristionchus), Panagrolaimidae (Panagrolaimus) and Cephalobidae (Cephalobus). A first diagnosis can be performed using a compound microscope, based on mouth morphology (Figure 3). There is, to date, no easy-to-use identification key for soil nematodes. The best way to identify the genus or the species is to compare with published drawings. For a detailed guide of soil nematodes, see Andrassy (1983, or 1984, in German); for the Rhabditidae, see Sudhaus and Fitch (2001); for the Caenorhabditis species, see Sudhaus and Kiontke (1996). Note that soil nematode diversity is huge and that most species are still undescribed. We detail below how to identify C. elegans and closely related Caenorhabditis species.
|
Figure 3. Schematic drawings of the characteristic buccal apparatus of different soil nematode families. Redrawn from Andrassy (1984). Note the characteristic long and straight stoma of Rhabditidae, the well-developed teeth at the base of a short stoma of Diplogastridae, the well-sclerotized stoma lining in Panagrolaimidae and the stoma lining divided in several elements of Cephalobidae.
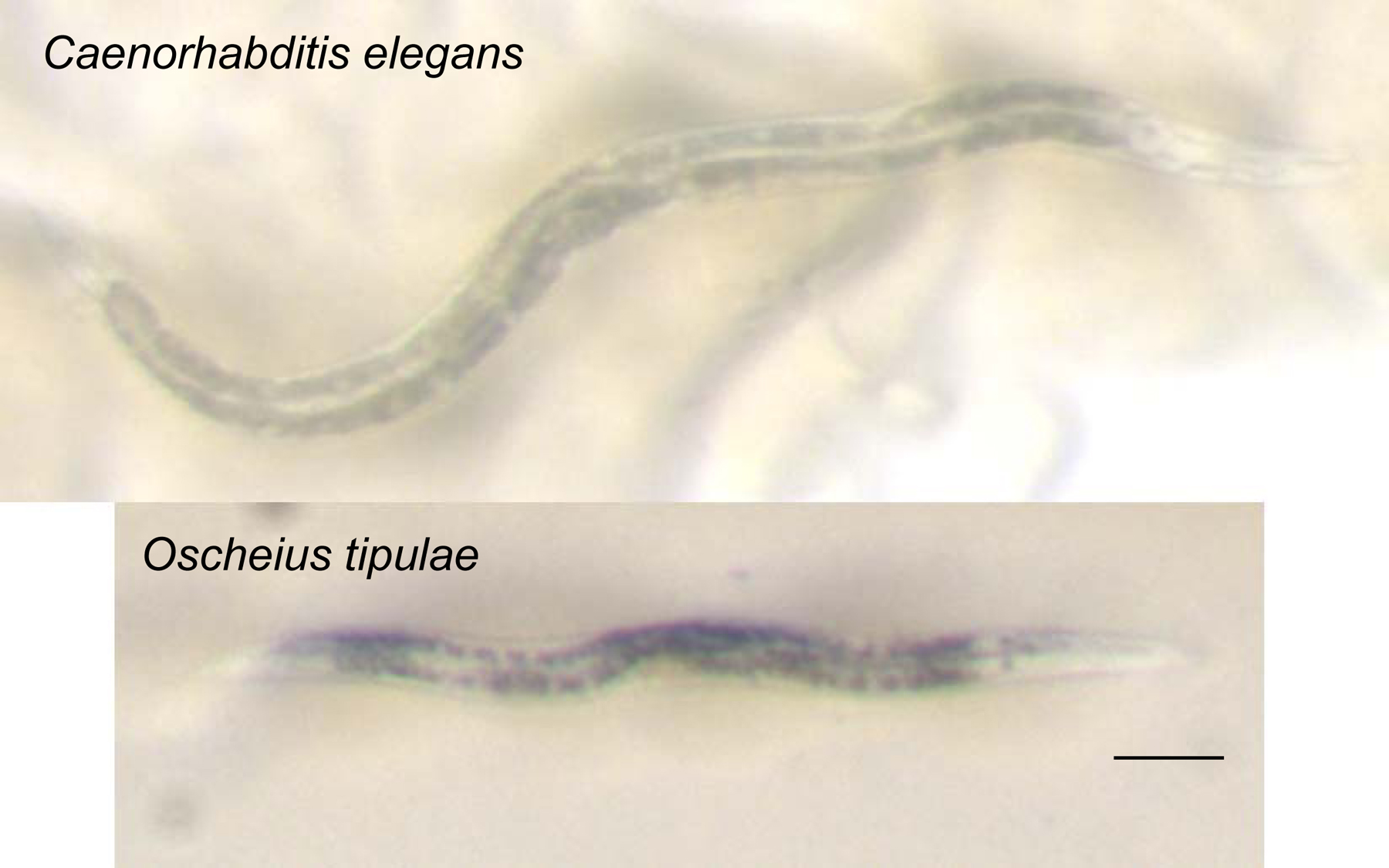
Here are some tips to identify Caenorhabditis under the dissecting microscope: Caenorhabditis hermaphrodites/females move relatively fast and elegantly, have a pointy tail, which folds back on itself when the worm is moving backwards. Most characteristically, the gut has a homogeneous light brown color under transmitted illumination and displays few large cell nuclei (which appear white). By contrast, most other nematodes have a greyer and darker gut, often with an antero-posterior asymmetry in pigment intensity, and with a large number of small or barely visible cell nuclei (Figure 4). The Caenorhabditis vulva is located towards the middle of the animal along the antero-posterior axis. With proper illumination, it is possible to see the second pharyngeal bulb (Figure 5).
|
Figure 4. Photographs of adult hermaphrodites of Caenorhabditis elegans (top) and Oscheius tipulae (bottom). Note the light brown color of the intestinal cell contents of Caenorhabditis compared to the darker color of the intestine of many other soil nematodes, including the common Oscheius tipulae. In addition, the few large intestinal cell nuclei of C. elegans are readily visible: compare them to the smaller nuclei of Oscheius tipulae. Notice also the smaller size of O. tipulae. Bar: 100 μm.
|
Figure 5. Drawings of Caenorhabditis elegans and Oscheius tipulae head and tail regions. Note the short rectum and very discernible second pharyngeal bulb (arrow) of C. elegans compared to the distinctive elongated rectum (arrow) and simple widening of the pharynx of O. tipulae.
In rich soil or in a compost sample, Caenorhabditis animals are mostly found in the dauer stage, which makes them difficult to recognize by the above criteria. By experience, the longest dauers of compost samples are often Caenorhabditis, whereas the smaller ones are of the Oscheius genus. If one waits for a week for the culture to bloom, Caenorhabditis may appear in the form of waving dauers (some other species also adopt this behavior, but not Oscheius).
When the worms are mounted and observed under a compound microscope, additional traits that characterize Caenorhabditis are its mouth shape (Rhabditidae; Figure 3) and the wide cytoplasmic rachis (see Figure 1 in Transformation and microinjection) in the center of the two-armed (didelphic) hermaphrodite/female gonad. Oscheius species can be best recognized by their long rectum (Figure 5).
Most C. elegans isolates are sensitive to RNAi by feeding. Sivasundar and Hey (2005) developed a screen based on this feature: worms extracted from a soil sample are transferred onto a plate seeded with bacteria expressing dsRNA against unc-22/twitchin (see Reverse genetics). C. elegans individuals can be recognized within a few days by their twitching behavior, while other nematode species will be unaffected, because most are probably insensitive to RNAi by feeding, and moreover, the degree of gene similarity is too low for the inactivation of the orthologous gene to occur. There is however a caveat, as some natural strains of C. elegans may be resistant to RNAi by feeding (Sivasundar and Hey, 2005).
Sequencing of the 18S ribosomal RNA (small subunit) gene with nematode-specific primers (Floyd et al., 2002; Haber et al., 2005) and blasting it against databases is another way to identify a species, or at least the group to which it belongs. Possible primers are:
SSU18A (5′-AAAGATTAAGCCATGCATG-3′)
and SSU26R (5′-CATTCTTGGCAAATGCTTTCG-3′),
which amplify a 1 kb fragment of 18S rDNA in rhabditid nematodes (Floyd et al., 2002), or
RHAB1350F (5′-TACAATGGAAGGCAGCAGGC-3′)
and RHAB1868R (5′-CCTCTGACTTTCGTTCTTGATTAA-3′),
which amplify a fragment of about 500 bp (Haber et al., 2005).
In order to discriminate between closely related Caenorhabditis species (especially the hermaphroditic species C. elegans and C. briggsae), one can perform a PCR using species-specific oligonucleotides (Barrière and Félix, 2005). For example, the following species-specific PCR primers in the glp-1 gene amplify different fragment sizes in C. elegans, C. briggsae and C. remanei. PCR can be performed using all 6 primers in the same reaction (annealing T° of 60°C).
C. elegans: CCGCTTGGATTCTATGGATTG and CTCTCCTTGCTCCGGGATTG, product: 208 bp.
C. briggsae: GAACCTGCGAGTGCATGTAC and CCGTCTGCAAACGAACGGGC, product: 302 bp.
C. remanei: CAACGGAGGTATCTGCTCAG and CCGCCGTCAAATTTGCATTC, product: 391 bp.
Different Oscheius species of the Oscheius tipulae group can be distinguished using a PCR-RFLP test (Félix et al., 2001).
Different Pristionchus species are distinguished by sequencing of 18S RNA (Herrmann et al., 2006).
Species identity can be confirmed by crossing several virgin (L4 stage) females or hermaphrodites (all progeny of a single individual) with males of identified laboratory strains (about 5 animals of each sex are placed onto a culture plate). In hermaphroditic species, successful crosses can be scored by the occurrence of a high proportion of male progeny; in female-male species, the presence of (fertile) progeny is a sufficient indicator. Crosses between some closely related species (such as C. remanei and C. briggsae; Baird, 2002) may yield a few (sterile) F1 progeny, but most of the eggs that are laid in these crosses never hatch. No progeny of C. elegans with any other species has been reported so far. Strains of many species can be obtained either from the CGC (http://biosci.umn.edu/CGC/Strains/strains.htm), the NYU Rhabditidae collection (http://www.nyu.edu/projects/ fitch/WSRN/) or our collection (http://www2.ijm.jussieu.fr/worms/).
A refined method is to use males of laboratory strains that express dominant markers (e.g., GFP). In this case, cross-progeny are identified by the presence of the marker.
The organization and timecourse of a nematode isolation experiment depend on its goal:
If the goal is to isolate a specific species, it may be best recognized once in the adult stage and may be easier to spot after one generation on the plate. Some species are resistant to harsh conditions and can be isolated by plating the sample several weeks/months after its collection. Caenorhabditis can be recovered after up to 10 days of sample collection, yet most likely with a much reduced efficiency compared to fresh samples.
If the goal is to obtain an accurate representation of genetic diversity of a natural population, one needs to isolate the individuals that were present at the time of sampling, that is within a single generation (obviously, the time between sample isolation and plating must be reduced to the minimum).
If the goal is to study wild worms and directly determine their number, sex, stage, associated bacteria, etc., the worms need to be isolated within minutes to a few hours after sample collection.
As mentioned above, Caenorhabditis and Oscheius are easily spotted as dauers on the first day of plating, or as L4s/young adults on the second day. Individuals of Pristionchus species are best recognized after two days by their characteristic ‘Dumpy’ appearance and the strong longitudinal ridges on the adult cuticle. Slow moving species that only become apparent after several days are cephalobs (these are also the major group of nematodes found in samples from dry soil) and Bunonema (tiny worms with a left-right asymmetric cuticle). Finally, when the plate is mostly depleted of bacteria and full of nematodes (6–10 days), dauers appear again, some of which adopt a waving behavior and may belong to the Caenorhabditis genus.
Be aware that some nematodes that can be found in soil samples such as Halicephalobus (Shadduck et al., 1979) or Pelodera (Ginsburg et al., 1984; Tanaka et al., 2004) have been described to be potentially harmful to humans. However, the major danger you may encounter is to contaminate your lab with wild mites, either nematophagous ones, swallowing worms like spaghettis, or bacterivorous ones, competing with nematodes and messing up cultures by transferring eggs and larvaes between plates. Good practice is keeping your sampling plates apart from your culture plates, and wrapping them with tight Parafilm. Mites can be killed by heating plates/incubators to 50–60°C for a few hours; alternatively, dichlorobenzene moth balls have been reported to kill mites but not nematodes (K. Kiontke, personal communication).
Some species of the Rhabditidae, Diplogastridae, Panagrolaimidae and Cephalobidae families can be grown on standard NGM plates seeded with OP50 (see Maintenance of C. elegans); however, NGM is too rich for some other species, which prefer water plates (1% agar in water + 5 μg/ml cholesterol).
Some species of other families require special culturing media or conditions (P. DeLey, personal communication). Bunonema, Tylocephalus and Plectus can be best cultured on soil plates (1% agar + 1% soil + 5 μg/ml cholesterol), and Teratocephalus on water plates (1% agar in water + 5 μg/ml cholesterol). Add ca. 2 ml of M9 solution to the Plectus and Teratocephalus cultures (they like to swim). Keep cultures at 15–20°C, except for Bunonema, which grows best at 25°C.
Male-female species may be subject to inbreeding depression after a few generations of culturing in the lab. This is best avoided by taking a large number of worms (>10) at each transfer and/or culturing them on larger (90 mm diameter) culture plates.
Many species can be frozen using the C. elegans freezing protocol (see Maintenance of C. elegans). However, several species do not survive freezing using this method.
For species of the Mesorhabditis, Teratorhabditis, Pelodera, Poikilolaimus, Oscheius (Insectivora group) and Panagrolaimus genera, we have had more success using the Pristionchus protocol (using tap water in the freezing solution and PBS instead of M9 to resuspend the worms; R. Sommer, personal communication; or adding 1 mM CaCl2 in the freezing solution, André Pires da Silva, personal communication).
Finally, species of the Cephalobidae family are particularly drought-resistant and can be frozen by directly placing large chunks of a half-dried culture plate (slightly before it turns into an agar ‘chip’) into a tube at −80°C; for ‘revival’, H2O or M9 solution is added to the tube for a few hours and the tube contents are then transferred to a new culture plate.
We thank Asher Cutter, Karin Kiontke and two anonymous reviewers for helpful comments.
Andrassy, I. (1983). Taxonomic review of the suborder Rhabditina (Nematoda: Secernentea). Paris: Orstom.
Andrassy, I. (1984). Klasse Nematoda (Ordnungen Monhysterida, Desmoscolecida, Araeolainzida, Chromadorida, Rhabditida). Stuttgart: Gustav Fischer Verlag.
Baird, S.E. (2002). Haldane's rule by sexual transformation in Caenorhabditis. Genetics 161, 1349–1353. Abstract
Barrière, A., and Félix, M.A. (2005). High local genetic diversity and low outcrossing rate in Caenorhabditis elegans natural populations. Curr. Biol. 15, 1176–1184. Abstract Article
Cutter, A.D. (2006). Nucleotide polymorphism and linkage disequilibrium in wild populations of the partial selfer Caenorhabditis elegans. Genetics 172, 171–184. Abstract Article
De Ley, P., and Mundo-Ocampo, M. (2004). Cultivation of Nematodes. In: Nematology-Advances and Perspectives. Z. Chen, S. Chen, and D.W. Dickson, eds., CABI publishing, pp. 541–619.
Félix, M.-A., Vierstraete, A., and Vanfleteren, J. (2001). Three biological species related to Rhabditis (Oscheius) pseudodolichura Körner in Osche, 1952. J. Nematol. 33, 104–109.
Floyd, R., Abebe, E., Papert, A., and Blaxter, M. (2002). Molecular barcodes for soil nematode identification. Mol. Ecol. 11, 839–850. Abstract Article
Ginsburg, B., Beaver, P.C., Wilson, E.R., and Whitley, R.J. (1984). Dermatitis due to larvae of a soil nematode, Pelodera strongyloides. Pediatr. Dermatol. 2, 33–37. Abstract
Haber, M., Schungel, M., Putz, A., Muller, S., Hasert, B., and Schulenburg, H. (2005). Evolutionary history of Caenorhabditis elegans inferred from microsatellites: evidence for spatial and temporal genetic differentiation and the occurrence of outbreeding. Mol. Biol. Evol. 22, 160–173. Abstract Article
Herrmann, M., Mayer, W.E., and Sommer, R.J. (2006). Nematodes of the genus Pristionchus are closely associated with scarab beetles and the colorado potato beetle in Western Europe. Zoology 109, 96–108. Abstract Article
Hooper, D.J. (1986). Extraction of free-living stages from soil. In: Laboratory methods for work with plant and soil nematodes. J.F. Southey, ed., London: Ministry of Agriculture, Fisheries and Food, pp. 5–30.
Kiontke, K. (1997). Description of Rhabditis (Caenorhabditis) drosophilae n. sp. and R. (C.) sonorae n. sp. (Nematoda: Rhabditida) from saguaro cactus rot in Arizona. Fundam. Appl. Nematol. 20, 305–315.
Shadduck, J.A., Ubelaker, J., and Telford, V.Q. (1979). Micronema deletrix meningoencephalitis in an adult man. Am. J. Clin. Pathol. 72, 640–643. Abstract
Sivasundar, A., and Hey, J. (2005). Sampling from natural populations with RNAi reveals high outcrossing and population structure in Caenorhabditis elegans. Curr. Biol. 15, 1598–1602. Abstract Article
Sudhaus, W., and Fitch, D. (2001). Comparative studies on the phylogeny and systematics of the Rhabditidae (Nematoda). J. Nematol. 33, 1–70.
*Edited by Ralf J. Sommer. WormMethods editor, Victor Ambros. Last revised May 10, 2006. Published July 17, 2006. This chapter should be cited as: Barrière, A. and Félix, M.-A. Isolation of C. elegans and related nematodes (July 17, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.115.1, http://www.wormbook.org.
Copyright: © 2006 Antoine Barrière and Marie-Anne Félix. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: felix@ijm.jussieu.fr
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.